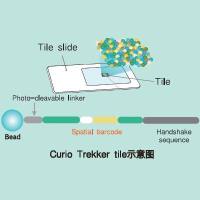
Minisatellite Variant Repeat Unit Mapping Using PCR (MVR-PCR)
互联网
627
Since the discovery of DNA fingerprinting ten years ago (1 ), the direct analysis of DNA variation has been successfully used in a wide range of applications and has become an important tool in forensic science (2 ,3 ). Most DNA typing systems assay DNA fragment length at repeated loci showing tandem repeats, for example, minisatellites and microsatellites, which exhibit allelic variation in tandem-repeat copy number (4 -10 ). However, despite the great discriminatory power and sensitivity of this approach, it has inherent limitations that have prevented its full potential from being realized. The most informative loci, hypervariable minisatellites, have quasicontinous allele-length distributions, with many more alleles than can be resolved by agarosegel electrophoresis, making unequivocal identification of individual alleles impossible (3 ,6 ,11 -16 ). This has led to protracted arguments over the definition of match criteria and the extent to which allele-size estimates are prone to error (17 -19 ). This problem can, in principle, be overcome by using microsatellites, which have fewer and smaller alleles that can be precisely sized on sequencing gels (9 ,10 ). Although microsatellites have now been widely adopted in the construction of investigative forensic databases, these loci also have attendant drawbacks, the most important being restricted allelic variability, as compared to hypervariable minisatellites.