


亚群专一性转录因子和Thl/Th2分化的表观遗传学因素 在细胞因子受体介导CD4丁亚群分化的信号转导中,参与的转录因子有两类是STAT家族,家族中个成员发挥作用取决于不同细胞因子及其配接的受体;二是亚群专一性转录因子,对Thl亚群主要是T-bet,对Th2亚群主要是GATA-3,对Thl7主要是RORyt。从图9-15可以看出,这些转录因子有三个特点,一是编码基因的激活早于Thl和Th2分 ...
记忆性T细胞的产生是一个由抗原启动的程序化过程 淋巴细胞介导的适应性免疫应答,就淋巴细胞的命运而言,经历了抗原特异性克隆的增殖、收缩和记忆三个时相。增殖指淋巴细胞的激活和发生克隆扩增;收缩指行使免疫效应后发生效应细胞的凋亡;记忆指记忆性淋巴细胞的分化和维持。三者的交替在不同程度上受到程序化的调控。其中的决定因素是抗原刺激的类别、强度和持续时间.记忆性T细胞的发育 记忆性T细胞的发育 ...
多因素制约CD4T亚群的分化 抗原类型、剂量、引入途径可制约亚群的分化。用不同剂量的病毒诱导新生小鼠产生抗病毒免疫应答。如果体内注入低剂量的病毒(0.3个噬斑形成单位),可以产生强有力的抗病毒效应,因为可诱导以IFN-7为主的Thl型细胞免疫。大剂量输入病毒(1 000个噬斑形成单位)后,主要诱导Th2型应答,所产生的Ⅱ/-4虽也有抗病毒效果.但不如前者。不同低剂量病毒感染诱导新生小鼠产生 ...
超抗原对T细胞的激活 超抗原是多种病原微生物如细菌、支原体和病毒的产物。大部分外源性超抗原来自细菌,常见的为金葡菌肠毒素(staphylococcal enterotoxln,SE)和毒性休克综合征毒素(toxic shock syndrome toxin-1,TSST―1)。病毒性超抗原为内源性,小鼠中较为常见,如小鼠乳腺肿瘤病毒(MMTV)。人体中T细胞对狂犬病毒和EB病毒的反应认为也 ...
记忆性T细胞长期维持的机制 患有烈性传染病并最终康复者,保护性免疫的持续时间可达60~?o年,有的终身免疫。疫苗接种后(例如天花疫苗),特异性抗体及记忆性细胞也据称可维持数十年。这些都提示,抗原特异性记忆细胞在体内可以长期存在。其中的机制尚未完全阐明,现有的解释包括两个方面。 1.残存抗原和交叉反应抗原的刺激 某些抗原(包括完整的病毒颗粒)可以以免疫复合物的形式在免疫组织的特定部位 ...

记忆性T细胞的异质性及其亚群 第一章提到,免疫记忆性有保护性记忆和反应性记忆之分。对T细胞应答,前者由效应性记忆T细胞(TEM)介导,后者由中枢性记忆T细胞(TCM)介导。TEM通常迁移至外周炎症组织,在二次应答中行使速发性效应功能;TEM则定居在外周淋巴器官的T细胞区,不直接发挥效应作用,但可在抗原再次刺激时,重新分化成为效应细胞,参与记忆性应答。 实际上,记忆性T细胞还划分成C ...
超抗原与TCR和MHC分子的结合 蛋白质抗原必须经过加工和提呈才能被T细胞识别,然而超抗原在激活T细胞之前无须加工和处理,因而具有两个不同点:一是激活T细胞的超抗原为完整的抗原分子而不是抗原肽;二是超抗原一般不进入MHC分子的抗原结合槽。这两点决定了超抗原和TCR/MHC发生相互作用时构成的三元体。与经典形式的三元体不同。超抗原分子位于TCR和MHC分子的外侧,一端和TCR日链相连,另一端 ...
初始B细胞的特性及其激活途径 从中枢免疫器官进入外周的B细胞尚未完全成熟,虽表达高水平的mlgM和低水平的mlgD,但只能存活几天。少部分B细胞可进入外周免疫器官的淋巴滤泡,成为成熟B细胞,半活期可达3~8周,并进入淋巴细胞再循环。决定其发育成熟为较长寿B细胞的因素,包括抗原的激发、信号转导、特定趋化因子如CXCLl3,以及B细胞激活因子(BAFF)及其受体BAFF-R等。 在外周 ...
不同的超抗原取用TCR声链的不同片段 人体TCRBV基因(又称Vp编码基因)共有64个片段而分成22个结构不同的家族。有意义的是.超抗原往往专一性地和某些家族的片段结合,即选择性地激活携带有这些TCR卢链的T细胞.由此构成超抗原对TCRBV基因片段取用(usage)的非随机性。表9―6列举了一些常见的细菌性超抗原对26个TCR9链家族的识别和取用特点。可以看出,B型和C2型金葡菌肠毒素(S ...
B细胞的抗原识别结构辅助受体和辅佐分子 在成熟B细胞表面,CDl9、CD21与CD81以非共价键组成B细胞活化的辅助受体复合物(CO-receptor complex)。它们所起的作用类似于T细胞表面的辅助受体CD4或CD8。 (1)CD21:又称Ⅱ型补体受体(CR2),为单链工型跨膜糖蛋白,分子质量为145kDa,胞外区由15个短同源重复序列(SCR)组成。配体是补体C3的裂解片 ...
BCR Igα/Igβ复合物 B细胞抗原受体(BCR)为膜型免疫球蛋白(mlg)。初始B细胞的BCR为mlgM和mlgD,活化和记忆B细胞则关闭mlgD的表达。一个B细胞可表达104一l05个mlg分子。 mlg与抗体在蛋白结构上基本相同,远膜端为分列的两个抗原结合部位,不同之处是多了跨膜区和极短的胞内部分。人mlg的跨膜区有两个高度保守的序列TAST和YSTTVT。前者与Ig从内 ...
B细胞抗原结合导致的受体交联启动信号传导 B细胞识别抗原后所启动的信号转导与T细胞十分相似,首先通过受体分子的交联成簇启动信号的跨膜传导,但两者启用的机制不同,因为B细胞可以识别完整的、具有多个表位的抗原分子而不是经由MHC分子提呈的抗原片段。抗原分子本身就可以通过表位与多个BCR结合,或如前所述借助抗原-补体复合结构同时结合BCR及CD21分子(BCR-Ag-C3dg-CD21),使B细 ...
B细胞的抗原识别信号转导的启动 静止状态时,Src家族的PTK如Fyn、Blk和Lyn通过酰基化与膜脂结合而附着于细胞膜内侧,因成簇和相互磷酸化而激活后,首先使结合于BCR的Iga/Igp分子胞内段上ITAM中的酪氨酸残基发生磷酸化,磷酸化的ITAM招募各种带有SH2结构域的蛋白质分子,首当其冲的是带有两个SH2结构域的Syk-PTK分子。Syk与T细胞中的ZAP-70一样,都属胞质中游离 ...
B细胞激活信号的胞内传违 膜结合的衔接蛋白是连接淋巴细胞激活信号胞内传递的重要成分,T细胞中为I-AT.B细胞中称为B细胞连接蛋白(Bcelllinkerprotein,BLNK)。前面提到,游离于胞质中的Syk-PTK被招募并激活后,其作用的靶分子之一,即是含SH2结构域的BLNK,并通过BI。NK构筑的“平台”,吸引并进一步激活多种信号分子包括蛋白激酶和其他衔接蛋白,Tec-PTK家族 ...
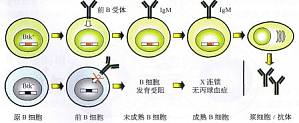
Bruton酪氨酸激酶与X性联无丙球血症 X性联无丙种球蛋白血症(X-1洫kedagammagl。bulinemia,XLA)是临床上首次确定的体液免疫缺陷病,由Brutor于1952年报道,故又称Bruton病。XLA作为常见的原发性免疫缺陷病之一,特点为血清中各类免疫球蛋白缺乏,或只能测得微量的抗体。外周血和淋巴组织中B细胞减少或完全缺乏,淋巴结中无生发中心和浆细胞。本病起病早,在出生 ...
T、B细胞相互作用与B细胞的增殖分化 B细胞对蛋白质抗原的应答及抗体的产生需要T细胞的协助,即要求Th2细胞提供B细胞激活和分化的第二信这一协助发生于T-B细胞间的相互作用,包括两个方面:B细胞从BCR获取抗原识别信号之后作为APC对Th2的激活;以及Th2表达CD40L和分泌细胞因子,协助B细胞进一步分化。 B细胞增殖分化有关的细胞因子 B细胞接受足够强度的双信号后从Go ...
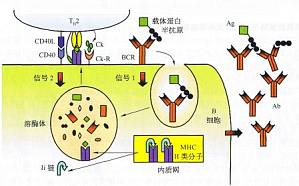
B细胞的抗原提呈作用 B细胞可通过BCR结合抗原并发生胞吞(cndocytosis)摄人BCR-Ag复合物。在抗原加工区室CⅡV中,抗原被降解成肽段,然后抗原肽与进入区室的MHCⅡ类分子结合,表达于B细胞表面并提呈给CD4T细胞。需要指出的是,B细胞以其BCR识别的抗原表位不同于它提呈的供T细胞识别的表位,两者分别来自半抗原和与之结合的载体蛋白(图10-6),也可以来自同一抗原分子的B表位 ...
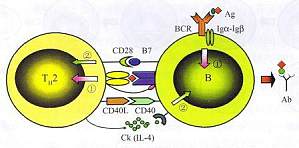
T、B细胞间的相互作用 1.T、B细胞相互作用的过程 T细胞为Th2亚群,而B细胞既是Th细胞辅助的对象,又是T细胞活化的APC。图10-7表明,这一相互合作包括一系列过程:①抗原以其B表位(绿色)与BCR结合传递B细胞活化的第一信号,使其C1340与细胞因子受体的表达增加,并使B细胞加工提呈抗原和表达少量D7分子;②B细胞作为APC将带有T表位(红色)的抗原肽提呈给CD4T细胞,供T细 ...
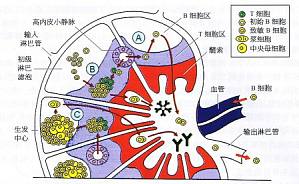
B细胞的增殖与分化在外周淋巴组织中进行。其中,包括B细胞进入B细胞区、识别抗原、与T细胞发生相互作用、出现增殖性原发灶、形成生发中心,并在生发中心中完成抗体亲和力成熟及类别转换,最终形成浆细胞及记忆B细胞。 B细胞可以经过两种途径进入外周淋巴结,一是经由输入淋巴管,二是穿越位于淋巴结中的HEV。下面将要提到,HEV位于T细胞区,该区中的基质细胞、HEV ...
生发中心的发育与结构 生发中心主要由B细胞组成,其中也有部分(大约10%)抗原特异性T细胞(有利于继续为B细胞分化提供辅助)及Mφ。在光镜下,由淋巴结的内层向外(图中为从下到上),生发中心的结构依次为暗区(darkzone)、亮区(1ightzone)和边缘区(mantlezone)。 生发中心的发育和形成经历三个阶段:①中央母细胞阶段:进入淋巴滤泡的B细胞每6~8h分裂一次,呈指数 ...
关于丁香通
公司信息
个人用户
企业机构


