


记忆性T细胞的异质性及其亚群 第一章提到,免疫记忆性有保护性记忆和反应性记忆之分。对T细胞应答,前者由效应性记忆T细胞(TEM)介导,后者由中枢性记忆T细胞(TCM)介导。TEM通常迁移至外周炎症组织,在二次应答中行使速发性效应功能;TEM则定居在外周淋巴器官的T细胞区,不直接发挥效应作用,但可在抗原再次刺激时,重新分化成为效应细胞,参与记忆性应答。 实际上,记忆性T细胞还划分成C ...

超抗原与TCR和MHC分子的结合 蛋白质抗原必须经过加工和提呈才能被T细胞识别,然而超抗原在激活T细胞之前无须加工和处理,因而具有两个不同点:一是激活T细胞的超抗原为完整的抗原分子而不是抗原肽;二是超抗原一般不进入MHC分子的抗原结合槽。这两点决定了超抗原和TCR/MHC发生相互作用时构成的三元体。与经典形式的三元体不同。超抗原分子位于TCR和MHC分子的外侧,一端和TCR日链相连,另一端 ...
初始B细胞的特性及其激活途径 从中枢免疫器官进入外周的B细胞尚未完全成熟,虽表达高水平的mlgM和低水平的mlgD,但只能存活几天。少部分B细胞可进入外周免疫器官的淋巴滤泡,成为成熟B细胞,半活期可达3~8周,并进入淋巴细胞再循环。决定其发育成熟为较长寿B细胞的因素,包括抗原的激发、信号转导、特定趋化因子如CXCLl3,以及B细胞激活因子(BAFF)及其受体BAFF-R等。 在外周 ...
不同的超抗原取用TCR声链的不同片段 人体TCRBV基因(又称Vp编码基因)共有64个片段而分成22个结构不同的家族。有意义的是.超抗原往往专一性地和某些家族的片段结合,即选择性地激活携带有这些TCR卢链的T细胞.由此构成超抗原对TCRBV基因片段取用(usage)的非随机性。表9―6列举了一些常见的细菌性超抗原对26个TCR9链家族的识别和取用特点。可以看出,B型和C2型金葡菌肠毒素(S ...
B细胞的抗原识别结构辅助受体和辅佐分子 在成熟B细胞表面,CDl9、CD21与CD81以非共价键组成B细胞活化的辅助受体复合物(CO-receptor complex)。它们所起的作用类似于T细胞表面的辅助受体CD4或CD8。 (1)CD21:又称Ⅱ型补体受体(CR2),为单链工型跨膜糖蛋白,分子质量为145kDa,胞外区由15个短同源重复序列(SCR)组成。配体是补体C3的裂解片 ...
BCR Igα/Igβ复合物 B细胞抗原受体(BCR)为膜型免疫球蛋白(mlg)。初始B细胞的BCR为mlgM和mlgD,活化和记忆B细胞则关闭mlgD的表达。一个B细胞可表达104一l05个mlg分子。 mlg与抗体在蛋白结构上基本相同,远膜端为分列的两个抗原结合部位,不同之处是多了跨膜区和极短的胞内部分。人mlg的跨膜区有两个高度保守的序列TAST和YSTTVT。前者与Ig从内 ...
B细胞抗原结合导致的受体交联启动信号传导 B细胞识别抗原后所启动的信号转导与T细胞十分相似,首先通过受体分子的交联成簇启动信号的跨膜传导,但两者启用的机制不同,因为B细胞可以识别完整的、具有多个表位的抗原分子而不是经由MHC分子提呈的抗原片段。抗原分子本身就可以通过表位与多个BCR结合,或如前所述借助抗原-补体复合结构同时结合BCR及CD21分子(BCR-Ag-C3dg-CD21),使B细 ...
生发中心的发育与结构 生发中心主要由B细胞组成,其中也有部分(大约10%)抗原特异性T细胞(有利于继续为B细胞分化提供辅助)及Mφ。在光镜下,由淋巴结的内层向外(图中为从下到上),生发中心的结构依次为暗区(darkzone)、亮区(1ightzone)和边缘区(mantlezone)。 生发中心的发育和形成经历三个阶段:①中央母细胞阶段:进入淋巴滤泡的B细胞每6~8h分裂一次,呈指数 ...
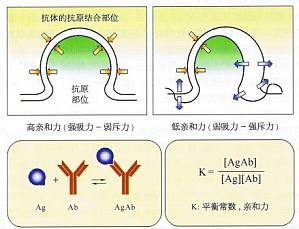
体细胞高频突变与抗体的亲和力成熟 抗体亲和力描述的是抗原表位与抗体分子抗原结合部位之间的互补情况,体现两种分子间吸引力与排斥力间的平衡与消长。亲和力可以用抗原-抗体复合物的形成量与游离抗原加游离抗体量的比值作定量评估,即计算两者间的平衡常数。抗原-抗体结合的亲和力及其测定原理图上方表明亲和力高低由分子间吸力和斥力间的消长所决定;下方为定量评估亲和力的平衡常数计算原理。 生发中心微 ...
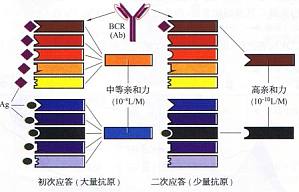
抗原对受体结构多样性B细胞克隆的选择 抗原对带有结构各异B(:R的B细胞克隆进行选择是亲和力成熟的关键。在初次应答时,大量抗原的出现,可使带有不同亲和力BCR的各种B细胞克隆被选择和激活,所产生的抗体,同时包括高、中、低亲和力,是一种混合物,其平均亲和力为中等。当大量抗原被清除,或二次应答仅有少量抗原出现时,该抗原会优先挑选高亲和力的3CR与之结合,仅仅使相应B细胞发生克隆扩增。其结果,该克 ...
抗体的类别转换 抗体主要履行三项功能:中和作用、调理作用和补体激活作用。对此,不同类别(class)的抗体各有侧重。而且,抗体的血清含量,穿越胎盘和向血管外弥散的能力、参与ADCC及诱导超敏反应的能力,也因类别不同而相互有别。显然,只有IgM一种类别的抗体是不行的。IgG类抗体在显示各种抗体功能,特别是抗细菌感染方面非常重要,并可穿越胎盘;而IgA对黏膜免疫、IgE对抗寄生虫感染实属必须。 ...
抗体类别转换的特点 类别转换不涉及抗体的抗原结合特异性,即不改变抗体的独特型(idiotype)。在分子水平,类别转换仅针对免疫球蛋白重链C区基因的重排,即通过C弘转换为CT、Co或CE而实现类别的改变。 类别转换主要出现在二次应答之后,由此产生的IgG随之发生亲和力成熟和含量的上升,表明两者基本上是同步的。抗体类别转换、亲和力成熟和抗体含量的提高发生在二次免疫应答A。类别转换( ...
影响抗体类别转换的因素 (1)抗原的性质:可溶性蛋白抗原(TD抗原)主要诱导人和小鼠产生IgGl;多糖类抗原易诱导IgM的产生,某些多糖对成年人诱导IgG2,对小鼠诱导IgG3;蠕虫类抗原易诱导IgE生成,可能与这些抗原易活化NKT细胞分泌IL-4有关。 (2)免疫途径与免疫佐剂:抗原免疫途径不同,产生的抗体类别也不相同。如口服抗原涉及黏膜免疫,产生IgA为主的抗体;而皮内、皮下免疫 ...
浆细胞的产生与抗体生成 在B细胞激活的次级时相经历亲和力成熟与类别转换的B细胞,最终分化成浆母细胞,后者再演变成分泌抗体的浆细胞。图10-17表明,浆细胞胞质中除了少量线粒体,几乎全部为合成抗体的糙面内质网所充斥,生动地表明,这是一个活的抗体生产“工场”。如果这一浆细胞来自生发中心,因为发生了类别转换,可以产生类别各异的抗体,定然不同于早期时相在T细胞区中出现的浆细胞,后者主要分泌IgM。 ...

二次抗体应答 Bm的表型和功能与初始B细胞有明显的区别。Bm长寿,不分裂或分裂非常慢,高表达IgM但不分泌抗体。Bm不易诱导耐受,遇到很低浓度的抗原即可被迅速激活,发生再次免疫应答,产生抗体的速度、性质、数量、亲和力、维持时间等都与初始B细胞介导的初次应答有很大的不同。再次应答的这些特点与记忆T细胞的存在,以及FIX;可以快速地结合免疫复合物有关。细胞对扣抗原的初次应答与再次免疫应答【生 ...
记忆性B细胞长期维持的机制 某些抗原(包括完整的病毒颗粒)可以以免疫复合物的形式在免疫组织的特定部位长期停留。其中起主要作用者为F1)(2。前面提到,该类细胞可借助表面的h受体及补体受体与“抗原-抗体”或“抗原-抗体-补体”复合物结合,在树突部分(细胞伪足)形成成串的颗粒状结构。称为免疫复合物覆盖小体(iccosome)。 FDC对这些复合物不作常规的吞噬和分解,而是将它们原封不动 ...
TI抗原的分类与主要特性 脂多糖及多糖类抗原能诱导无胸腺裸鼠或无T细胞动物产生抗体,称T细胞非依赖抗原(TI-Ag)。B细胞对TI抗原的抗体应答一般不出现二次回忆性反应,也没有抗体的亲和力成熟和类别转化。这意味着,TI抗原激发的抗体主要为IgM。 TI抗原因结构和作用机制不同,分为1型和2型。区别在于:①TI-1抗原主要是细菌细胞壁成分。其中的LPS属B细胞多克隆激活剂;而TI-2 ...
B细胞对TI抗原的应答 1.TI-1抗原诱导的抗体应答 LPS作为TI-1抗原的代表,引发的抗体应答可以有以下两种不同的机制。 (1)对B细胞多克隆的非特异性激活:见于抗原浓度高时,通过与B细胞表面LPS受体的结合,激活多克隆B细胞,产生低亲和力的IgM类抗体。此即LPS所具有的多克隆激活剂(polyclonal activator)功能,或称B细胞丝裂原(B cell mitog ...
CTL对靶细胞的杀伤 (一)CTL的分化成熟 CD8CTL在体内以非活化的前体细胞(CTL-P)形式存在,它必须经过抗原激活并在Th协同作用下才能分化发育为效应CTL。CTL的活化需要双信号。第一信号来自TCR特异性识别靶细胞膜上MHCI类分子-抗原肽复合物,并通过CD3分子参与抗原识别信号的传递。第二信号来自CTL细胞膜表面各种辅佐分子如CD28、CD2、LFA-1和靶细胞表明相 ...
CTL杀伤靶细胞细胞裂解性杀伤 (1)穿孔素:电镜下发现培养的CTL克隆在细胞内有电子致密物质。将这些颗粒分离出来后其本身就可介导靶细胞损伤。对这些颗粒进行分析后发现,其中含有高分子质量的糖蛋白,包括各种毒性细胞因子,如TNF-β、穿孔素(perforin,Pf)及一些蛋白酶如颗粒酶(granzymes,Gz)或片段酶(fragmentins)。这些酶都具有丝氨酸酯酶的活性。CTL-P通常 ...
关于丁香通
公司信息
个人用户
企业机构


