

- 询价
- 2025年07月16日


万千商家帮你免费找货
0 人在求购买到急需产品
- 详细信息
- 询价记录
- 技术资料
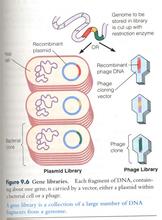
λ噬菌体
λ噬菌体是比细菌还小得多的微生物,和病毒侵犯真核细胞一样,噬菌体侵犯细菌,也可以认为它是细菌里的“寄生虫”。它本身是一种核蛋白,核心是一段DNA,结构上有一个蛋白质外壳和尾巴,尾巴上的微丝可以把噬菌体的DNA 注入细菌内。
λ噬菌体是一种双链DNA 噬菌体,基因组约为50kb+。线性λ噬菌体DNA 分子的两端各有12碱基的互补单链序列,是天然的粘性末端,称为COS 位点。
λ噬菌体生物学特性
λ噬菌体侵犯细菌时,只是DNA 侵入,然后利用细菌的酶来复制、转录、翻译。入侵E.coli后,可以按裂解或建立溶原两种生活方式生存和繁殖。
1、λ噬菌体在宿主内进行独立复制,在1 个菌体内生长出100 个左右的新噬菌体,并冲破(杀死)细菌释放出来,再度感染其它活菌,称为裂解。
2、λ噬菌体侵入菌体后,把自身DNA 加进细菌DNA 里(整合),作为细菌基因的一部分,随菌的细胞分裂而增殖,这种生活状态称为溶原。
溶菌(裂解)生长时在平皿上形成清亮的噬菌斑;溶源性生长。时在平皿上形成浑浊的噬菌斑。
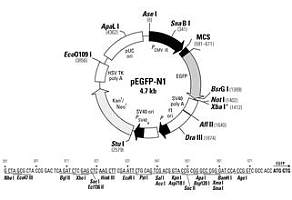
λ-DNA 载体的构建
1、缩短长度
野生型λ-DNA 包装的上限为51kb,本身长度为48.5kb,只有当插入的外源DNA 片段不大于2.5kb时,才能被包装成有感染力的噬菌体颗粒。因此缩短野生型λ-DNA 的长度,可以提高装载量。其实野生型λ-DNA 上约有40-50%的片段是复制和裂解所非必需的。
2、删除重复的酶切位点
野生型的λ-DNA 链上有5 个EcoRI 位点和7 个HindIII 位点,不利于重组操作,必须删除至1 - 2 个;同时,为了便于各种来源的DNA 片段的克隆,还需要增加一些单一的酶切位点;除了简单的切割外,还需要采用定点突变技术去除或增添酶位点。
3、加装选择标记
与质粒不同,野生型λ-DNA 上缺少合适的选择标记,因此加装选择标记是λ-DNA 克隆载体构建的重要内容。
λ-DNA 克隆载体上的选择标记主要有下列两类:
a、免疫功能类标记
加装选择标记imm434
imm434 基因编码一种阻止λ-噬菌体进入溶菌循环的阻遏物。含有完整标记基因的λ-载体进入受体细胞后,建立溶原状态,细菌生长缓慢,形成浑浊斑;当外源DNA 插入到标记基因中,基因灭活,λ-重组分子便进入溶菌循环,形成透明斑。
b、颜色反应类标记
加装选择标记lacZ
lacZ 基因编码β-半乳糖苷酶,能催化无色的X-gal 生成蓝色化合物。当外源基因插入到lacZ基因中,基因灭活,不能合成蓝色化合物;而空载体λ-DNA 则产生蓝色透明斑。
4、构建琥珀密码子的突变体
琥珀型突变(sup)是指由CAG(Gln)向UAG(stop)的突变。
肠杆菌的某些菌株含有特异性的校正基因,其编码产物校正tRNA 能专一性地纠正这一突变。将野生型λ-DNA 上D 和E 两个头部包装蛋白的基因中的CAG 密码子突变成UAG。当这种λ-DNA 进入一般的大肠杆菌菌株后,不能合成有活性的头部蛋白,也就不能被包装和裂解细菌,这样就可以阻止有害重组体的生物污染及扩散,而基因工程实验中使用的受体则是具有琥珀型突变体校正功能的菌株。
λ-DNA 载体的主要类型
插入型载体: Charon2、Charon6、λgt11
取代型载体:λEMBL4、λgtλc、λNM762、Charon40
λ-DNA 重组分子的体外包装
λ-DNA 重组分子需在体外人工包装成有感染力的噬菌体重组颗粒,方可高效导入受体细胞。
品化。这些包装蛋白通常分为相互互补的两部分:一部分缺少E 组份,另一部分则缺少D 组份。包装时,只有这两部分包装蛋白与重组λ-DNA 分子混合后,包装才能有效进行,任何一种蛋白包装液被重组λ-DNA 污染后,均不能被包装成有感染力的噬菌体颗粒,这也是基于安全而设计
λ-DNA 作为载体的优点
1、λ-DNA 可在体外包装成噬菌体颗粒,能高效转染大肠杆菌
2、λ-DNA 载体的装载能力为25 kb,远远大于质粒的装载量
3、重组λ-DNA 分子的筛选较为方便
4、重组λ-DNA 分子的提取较为简便
5、λ-DNA 载体适合克隆和扩增外源DNA 片段,但不适合表达外源基因
风险提示:丁香通仅作为第三方平台,为商家信息发布提供平台空间。用户咨询产品时请注意保护个人信息及财产安全,合理判断,谨慎选购商品,商家和用户对交易行为负责。对于医疗器械类产品,请先查证核实企业经营资质和医疗器械产品注册证情况。
- 作者
- 内容
- 询问日期
 技术资料
技术资料暂无技术资料 索取技术资料
λ噬菌体载体构建
询价
