
IL-35
简介:
IL-35 是 2002 年在第 13 届免疫学国际会议上被正式命名的一种新的抗炎 / 免疫抑制细胞因子 。2007 年, Vignali 和 Liew 2 个小组分别发表了有关 IL-35 生物学功能的论文 ,使其成为近期免疫研究的新热点。 IL-35 是由 EB 病毒诱导基因 3(Epstein-
Barr virus-induced gene 3 , EBI3)蛋白和 IL-12p35(IL-12A)亚基组成的异源二聚体,与 IL-12 、 IL-23 和 IL-27 共同组成了 IL-12 细胞因子家族,在 T 细胞增殖活化和细胞因子产生方面发挥重要的调节作用。研究证实, IL-35 主要由调节性 T 细胞(regulatory T cells , Treg)分泌,是 Treg 发挥免疫负调控的主要细胞因子之一 ,在多种疾病(如实验性结肠炎、胶原诱导性关节炎、自身免疫性脱髓鞘炎、慢性肝炎、糖尿病、肿瘤等) 中参与炎症的免疫调节,与疾病的发生、发展密切相关。 因此, IL-35 作为继转移生长因子 β(transforming growth factorβ , TGF-β) 和 IL-10 之后出现的新抗炎因子,有望成为治疗感染性疾病、自身免疫性疾病和肿瘤的靶点。
分子性质:
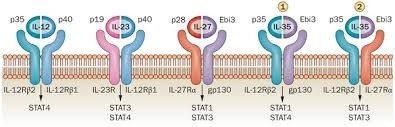
IL-35 具有 IL-12 细胞因子家族的典型特点,由 IL-12 的 α 链 IL-12A 亚基和 IL-27 的 β 链 EBI3 亚基组成的异源二聚体蛋白(图)。 人 IL-12A 亚基有 197 个氨基酸残基 , 含 7 个半胱氨酸和 3 个 N 糖基化位点,小鼠 IL-12A 有 193 个氨基酸残基,与人的 IL-12A 有 66% 同源性。 EBI3 亚基 1996 年首先发现于 EB 病毒(EBV)感染的 B 淋巴细胞培养上清中,与有 2 个 Ⅲ 型纤连蛋白结构域的 IL-12p40 亚基同源,是 IL-35 与 IL-27 共享亚基,属于造血细胞因子受体家族成员,由人类组织相容性抗原 (HLA)-G 提呈。编码 EBI3 蛋白的基因位于 19p13.3 ,邻近 EpoR 、非受体酪氨酸激酶 Hy1 、 Tyk2 及 CD23 基因,全长 1 161 bp ,编码 229 个氨基酸残基蛋白质, 相对分子质量约为 26 000 ,成熟 EBI3 蛋白由 209 个氨基酸残基组成,是一种可溶性细胞因子受体。
生物学活性:
研究发现, IL-35 2 个亚基的表达存在种属差异性 。在人类 IL-12A 亚基以极低浓度表达于脑、肠和肾组织, EBI3 亚基以低浓度表达于眼、 淋巴结和胰腺,但在人类胚胎期合胞体滋养层 2 个亚基同时高表达,表明 IL-35 可能是胎儿与母体之间的一种重要免疫耐受调节因子; 在鼠类 IL-12A 亚基和 EBI3 亚基则表现为高表达,并且在血液、骨髓、肝和胸腺均可检出 IL-12A 亚基, EBI3 亚基还高水平表达于鼠的淋巴结。 由此提示, IL-35 组成性表达于鼠的血液、骨髓、肝和胸腺等组织,而在人类组织则属于诱导性表达,进一步提示在不同物种中,分泌 IL-35 的细胞以及 IL-35 的分泌方式可能存在差别。
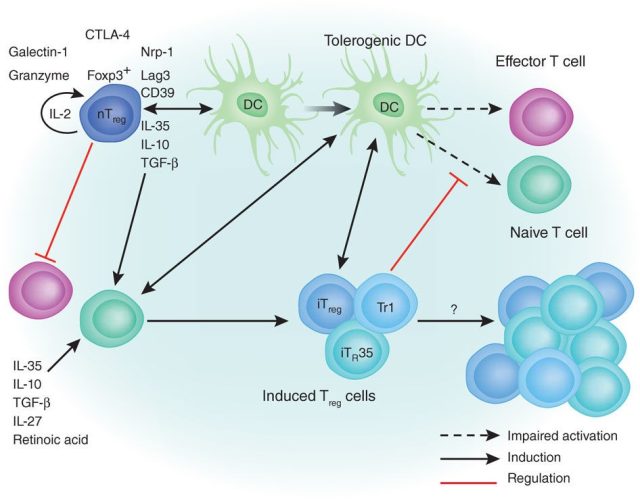
IL-35 分泌方面的研究目前主要集中在 Treg ,在鼠类可由未经活化的 Treg 分泌, 而在人类仅由活化的 Treg 分泌。 Treg 主要分为天然调节性 T 细胞(naturally occurring regulatory T cells ,nTreg)和诱导调节性 T 细胞(induced regulatory T cells , iTreg)2 大类。 Treg 主要通过分泌 TGF-β、IL-10 和 IL-35 3 种细胞因子而发挥免疫抑制作用, 而其中 TGF-β 是诱导 iTreg 分化的重要细胞因子,与 TGF-β 相似, IL-35也可诱导一种新型的 Treg 分化,称为 iTr35 ,其通过分泌 IL-35 起免疫抑制作用 。
另研究显示 ,在人类非 T 细胞系,如微血管内皮细胞、主动脉平滑肌细胞和上皮细胞,经促炎细胞因子 TNF-α 、 IFN-γ 和 IL-1β 刺激后 IL-35 表达上调。 然而诱导 IL-12A 亚基与 EBI3 亚基上调的细胞因子并不完全相同,提示 IL-35 的表达上调可能受双门道控制(double-gated ),原因是 2 个亚基受不同信号通路作用的结果。
IL-35 的受体分别是 IL-12Rβ2 和 gp130 , 二者可以组成异源二聚体,也可以各自组成同源二聚体。在缺少一条受体链的组织,传统 T 细胞对 IL-35 介导的免疫抑制表现敏感。因此推测,在不表达 IL-12Rβ2 的组织, IL-35 可能通过同源二聚体 gp130-gp130 传递活化信号 。 IL-35 的 2 个亚基分别与相应受体的不同部位结合,通过转录因子 STAT1 和 STAT4 传递转录信号。另外,有人推测 IL-35 可能拥有自独特的受体,详细机制有待于进一步研究探讨。
临床应用:
人类或鼠的初始 T 细胞经 IL-35 刺激后产生 iTr35 。iTr35 能分泌 IL-10 、TGF-β 和 IL-35 ,抑制 T 细胞增殖效应,有效阻止炎症的发生、发展,是诱导感染性耐受和促进 Treg 介导的肿瘤进展的关键因子。另一方面, IL-35 在体内外均可抑制 Th17 的分化及
IL-17 的产生,可明显降低炎症反应,因而与多种疾病密切相关。
1. IL-35 与感染性疾病 :慢性乙型肝炎患者外周血中 CD4 + T 细胞有 IL-12A 和 EBI3 的共表达,2 亚基可能以组成 IL-35 的形式存在,在慢性乙肝的免疫应答中发挥作用。
2. IL-35 与肿瘤 :与人类正常细胞系不同, IL-35 的 2 个亚基共表达在绝大多数人类肿瘤细胞系。在体外,肿瘤细胞上过表达的 IL-35 能有效降低肿瘤细胞活性,抑制肿瘤细胞生长,诱导细胞周期阻滞在 G 1 期, 通过上调 Fas 的表达、 下调细胞周期蛋白 D1(cyclin D1 )、生存素(survivin)和 B 细胞淋巴瘤 / 白血病 -2(Bcl-2)的表达增强肿瘤细胞凋亡的敏感性,介导强大的细胞凋亡效应 [19] ,为肿瘤的免疫治疗提供了新思路。
3. IL-35 与关节炎:IL-35 作为一个潜在治疗自身免疫性疾病的抑制性蛋白, 在不同类型关节炎中可能发挥不同的作用,其确切机制有待于进一步研究。
4. IL-35 与支气管哮喘:IL-35 在哮喘的控制和肺通气功能的改善方面发挥重要的调节作用,为哮喘的生物治疗提供了初步依据。
IL-35 作为 IL-12 细胞因子家族的新成员,已被大量研究证实是一种重要的抑制性细胞因子,在 Treg 细胞发挥免疫应答效应过程中起着重要的调控作用,与多种临床疾病密切相关。拮抗或促进 IL-35 的表达可能对维持机体免疫平衡具有重要意义,因而成为近来免疫学领域研究的热点问题之一。 然而,细胞因子之间及与细胞之间存在复杂的调节网络,IL-35 与其他细胞因子的关系及 Treg 细胞之间的密切联系仍有待于进一步研究, 其受体及信号传导途径目前仍不完全清楚,有待于深入探讨,以期为临床相关疾病的生物治疗提供理论依据。
-
 大肠杆菌高效表达外源蛋白的策略
大肠杆菌高效表达外源蛋白的策略大肠杆菌表达系统是发展最早、目前最为成熟的表达系统。因其遗传背景清楚、繁殖快、成本低、表达量高、表达产物容易纯化、稳定性好、抗污染能力强以及适用范围广等,是...
-
 His 融合蛋白纯化常见问题解答
His 融合蛋白纯化常见问题解答蛋白过镍柱纯化的原理:Ni-NTA 纯化介质纯化带有 His6-Tag 的融合蛋白是目前蛋白纯化中最常使用的一种方法。Ni 柱中的氯化镍或者硫酸镍可以与有 HIs(组蛋白)...
-
 GST 融合蛋白纯化常见问题指南
GST 融合蛋白纯化常见问题指南谷胱甘肽 S 转移酶(GST)是一个含有 211 个氨基酸的蛋白,通常将该蛋白加入到重组蛋白的末端以便对该重组蛋白进行纯化或检测。具有组氨酸的非融合蛋白会产生非特...
-
 DNA 甲基化修饰
DNA 甲基化修饰DNA 甲基化修饰作为一种重要的表观遗传修饰,能通过影响染色质结构、DNA 构想、稳定性以及蛋白质相互作用方式等,起到调控基因表达的作用。与多种肿瘤的发生、发展密...
-
 组织分离优化指南
组织分离优化指南尽管对于特定细胞种类的细胞分离步骤的优化,与细胞恢复的各种所需特性有关,但仍可以建立通用的优化指南。结合合理的实验设计方案,以下就细胞分离和酶的使用提出优化...
-
 如何 30 秒完成抗体标记?
如何 30 秒完成抗体标记?英国生物偶联技术专家级公司 InnovaBiosciences 为您提供一步法快速标记试剂盒!Perfect!Innova Biosciences 成立于 2002 年,是世界领先的生物偶联技术公司。...
-
 昆虫杆状病毒表达系统
昆虫杆状病毒表达系统昆虫杆状病毒表达系统 (baculovirus expression vector system, BEVS) 以昆虫杆状病毒为外源基因载体, 昆虫和昆虫细胞为受体的表达系统。昆虫细胞与哺乳动...