


亦称氨酰tRNA连接酶,氨基酸活化酶,是合成氨酰tRNA的酶。20种氨基酸,均有其相应的专一性的氨酰tRNA合成酶(ARS)中。一般认为它参与下列所示的两步反应,在ATP存在下使氨基酸活化,并与tRNA的CCA末端结合。 此酶能专一性地辨认氨基酸的侧链和tRNA。因为由于具有这样高专一性底物的酶,所以mRNA的遗传信息能准确无误地反映在蛋白质的氨基酸序列上。 ...

氨基酸的羧基与 tRNA的CCA末端腺苷的3′(或2′)羟基相结合的酯键。在蛋白质的生物合成中,它作为氨基酸转移到肽链的羧末端过程的中间物而发挥重要的作用。 ...
一种蛋白酶。属于肽链端解酶,可使氨基酸从多肽链的N-末端顺序逐个地游离出来。在许多生物中发现了各种性质的这种酶。具有代表性的是亮氨酸氨肽酶,特殊的有仅作用于N末端为脯氨酸的脯氨酸亚氨肽酶以及只作用于三肽的氨基三肽酶等。 ...
催化由NH3、CO2与2分子的ATP合成氨甲酰磷酸的不可逆反应的酶,EC2.7.2.5。存在于哺乳类的肝脏、蛙的肝脏、大肠杆菌等,分子量31.5万(蛙肝)。对于动物酶的活性,N-乙酰谷氨酸是必需的,经此酶生成的氨甲酰磷酸是生成精氨酸、尿素或合成嘧啶的素材。大肠杆菌的酶亦能利用谷酰胺作为氨基供体。此外在蕈类,如伞菌(Agaricus),存在着只利用谷酰胺作为氨基供体的酶(EC2.7.2.9)。 ...
为高能磷酸化合物之一,由氨和二氧化碳消耗2分子的ATP形成的(氨甲酰磷酸合成酶)。同时也已知道有利用谷酰胺作为氨基供体的酶的存在。其次此化合物可把氨甲酰基转移给尿素循环的鸟氨酸,形成瓜氨酸与磷酸。此外,与天冬氨酸反应可生成氨甲酰天冬氨酸。 ...
系乙酰胆碱衍生物。其作用类似乙酰胆碱,但不为胆碱酯酶分解。可用于治疗青光眼等。
催化氨基甲酸与ATP生成氨甲酰磷酸的反应酶EC2.7.2.2。存在于大肠杆菌、酵母、链孢霉属(Neu-rosPora)和豆类中。在此反应中生成的氨甲酰磷酸被用于精氨酸或氨甲酰天冬氨酸的合成。由于反应是可逆的,所以推测可由精氨酸代谢产生ATP。 ...

也称为转氨酶。是催化把α-氨基酸上的氨基转移给α-酮酸形成新的酮酸和氨基酸反应的酶类之总称是D.Needham(1927)在鸽胸肌中发现的氨基转移作用,后来A.E.Braunstein和M.G.Kritzmann等研究了此酶的性质,发现几乎在所有生物中都存在着这种酶。已知有谷氨酸氨基转移酶和天冬氨酸氨基转移酶等各种氨基酸特异的氨基转移酶,反应是可逆的,参与氨基酸的生物合成。以磷酸吡哆醛为辅酶。 ...
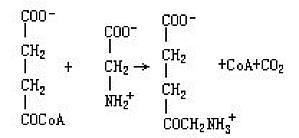
亦称δ-氨基酮戊酸合成酶。催化由琥珀酰CoA与甘氨酸生成δ-氨基-γ-酮戊酸反应的酶。EC2。3.1.37。在卟啉合成的中间阶段起作用。需要磷酸吡哆醛。存在于线粒体中,是代谢转换很快的酶。可为丙烯基异丙基乙酰脲(allylisopropy lacety-lurea)所诱导,还受血红素的反馈抑制。 ...
具有5-氨基-4-氧戊酸的结构,广泛分布于各种生物中。通常由琥珀酰CoA与甘氨酸经由α-氨基- β-酮己二酸生物合成的。2分子的δ-氨基-γ-酮戊酸脱水缩合,生成胆色素原。后者成为血红素、叶绿素、藻胆色素(phycobelin)、胆色素、维生素B12(氰钴胺素cyanocobalamin)等四吡咯化合物生物合成的中间产物。此外,还知有δ-氨基-γ-酮戊酸经氧化脱氨反应,生成α-酮戊二酸半缩醛 ...
缩写为ACC。为含于梨及苹果果汁中的非蛋白性质的氨基酸,为环状氨基酸的一种。植物体由蛋氨酸合成的乙烯的直接前体,在酶的作用下由S-腺苷酰蛋氨酸在活体内合成。许多植物组织都具有在氧的条件下使ACC分解生成乙烯的活性。 ...
含氨基糖的一组抗生素,链霉素为其代表。由链霉菌(Streptomyces)产生的其他的氨基糖苷抗生素有卡那霉素、新霉素及巴龙霉素等。都与30S核糖体亚单位相结合,引起m RNA之密码误读,从而抑制细菌的蛋白质活体合成。 ...
糖的羟基为氨基所取代的化合物的总称。作为生物体成分最常见的是葡糖胺和半乳糖胺,它是己糖的2位羟基为氨基所取代的化合物、神经氨酸,它是5位具有氨基的九碳糖,但从1位到3位具有丙酮酸的结构,如除去此部分则与2位具氨基的甘露糖胺的结构一致。与此不同,也有相当多的天然的2位以外的氨基糖,其大多数是作为微生物产生的核多糖、糖苷或抗生素的成分而存在的。例如作为糖构成成分的有大肠杆菌或色杆菌属(Chromo ...
为水解断开氨基而生成氨的酶的总称。对于腺嘌呤、腺苷、AMP、胞苷均各有其专一性的酶,而生成相应的OH基化合物。 ...
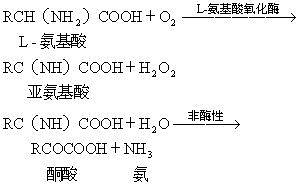
L-氨基酸氧化酶,存在于肾脏 肝脏 霉菌、蛇毒、细菌等中,为直接氧化L-氨基酸的需氧性脱氢酶。EC1.4.3.2.如果不存在过氧化氢酶,则以H2O2脱羧形成酮酸。 其辅酶是FMN或FAD。底物专一性广泛,能很好地氧化亮氨酸、甲硫氨酸等,但不作用于天冬氨酸、谷氨酸、甘氨酸等。但是,鸟类肝脏的酶对碱性氨基酸具有专一性。不过,由于活性低,因此动物的酶对氨基酸代谢是否有意义尚属疑问。但对L-α-羟酸 ...
催化脱去某种氨基酸的羧基,生成对应的胺(如下列括弧中所示)的裂解酶之总称:NH2CHRCOOH─→NH2CH2R+CO2。主要是在酸性培养基上发育的细菌中发现的,在动物,植物中虽然很少,但也可以找到。在细菌中已知有分别对应于赖氨酸(尸胺)、酪氨酸(酪胺)、精氨酸(鲱精胺)、鸟氨酸(腐胺)、谷氨酸(γ-氨基丁酸)等的专一性的脱羧酶,这些酶均以磷酸吡哆醛为辅酶。在动物组织中对谷氨酸(γ-氨基丁酸) ...
组成蛋白质的基本结构的多肽链中氨基酸残基的排列方式,即蛋白质的一级结构。特定的蛋白质各有其单独的氨基酸排列顺序,这是由对应的基因DNA(某种病毒则为RNA)中的核苷酸排列顺序所规定的。 ...
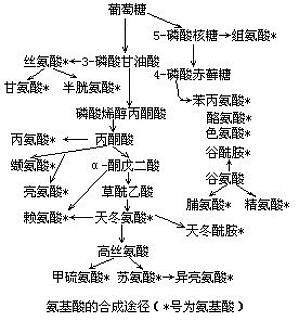
组成蛋白质的大部分氨基酸是以埃姆登-迈耶霍夫(Embden-Meyerhof)途径与柠檬酸循环的中间物为碳链骨架生物合成的。例外的是芳香族氨基酸、组氨酸,前者的生物合成与磷酸戊糖的中间物赤藓糖-4-磷酸有关,后者是由ATP与磷酸核糖焦磷酸合成的。微生物和植物能在体内合成所有的氨基酸,动物有一部分氨基酸不能在体内合成(必需氨基酸)。必需氨基酸一般由碳水化合物代谢的中间物,经多步反应(6步以上)而 ...
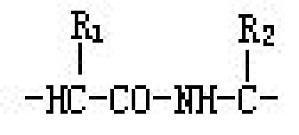
既含氨基(-NH2)又含羧基(-COOH)的有机化合物。氨基酸中还含有氨基的氢与分子中的其他部分发生取代而形成亚胺的环状化合物(亚氨基酸)。氨基与羧基结合在同一碳原子上的称为α-氨基酸。天然得到的氨基酸大部分是α-氨基酸(R-CHNH2-COOH),α-氨基酸相互间失水形成肽键连接(见图)的化合物为蛋白质或肽。由于氨基从α顺次向相邻的碳原子移动,因此被称之为β-,γ-,δ-氨基酸等,但并不存在 ...
组成蛋白质基本结构的多肽链两端,原则上是一端有一个氨基末端残基,另一端带有一个羧基末端残基。前者是将α氨基保持游离状态的氨基酸残基,后者是将α羧基保持游离状态的氨基酸残基。采用1-氟-2,4-二硝基苯的桑格(F.Sanger)的DNP法(DNP method),在历史上是很著名的。该法可作为鉴定或定量分析位于氨基末端的氨基酸残基的方法。也广泛采用将DNP法超微量化的dansyl法(用5-二甲氨 ...
关于丁香通
公司信息
个人用户
企业机构


