


荠的果形有三角形(AABB)和筒形(aabb)两种,三角形和筒形杂交,F1代的果形都是三角形,F2代只有两种表型三角形和筒形,比例为15:1,这也是9:3:3:1的变形。从此例可知,除 aabb以外,任何基因形,只要有一个A或B,表型都为三角形。A和B的作用相同,本身是完全显性,A和B互为完全显性,称等上位(isoelnstatic)或双显性上位(double dominant epistasis ...
将黑色的小鼠和白色小鼠杂交,F1代是黑色的,再将F1互交产生的F2有三种表型,黑色,棕色,白化,比例为9:3:4。产生的原因是形成黑色素的过程是一个连锁反应。当C、B基因都正常时,才能产生黑色素,当B基因发生突变而C正常时虽不能产生黑色素,但有棕色的中间产物形成。当C发生突变时,即使B基因正常也因缺乏中间产物而不能合成黑色素,反应停止在无色色素原这一步,所以小鼠呈白色。若C,B都发生突变小鼠同样呈 ...
香碗豆有紫花品系(PPCC)和白花品系(ppcc),杂交的F1代皆为紫花(PpCc)自交后F2代,只有紫花 (P_C_) 和白花 (P_ccC_ccppcc)两种表形,比例为9:7,当然也是9:3:3:1之比的变形。也就是说只有当C,P基因都正常时才开紫花。这是由于紫色素的合成是经过了一系列的连锁反应,无论是C基因还是P基因发生突变,此反应都不能完成,而中间产物是白色的 所以只有C_P_ 基因型呈 ...
显性上位产生3种表型,比例为12:;3:1,但产生这种比例的机制可能有不同的途径,一种是抑制基因所引起的。在植物中有一种可用于治疗心脏病的植物叫毛地黄(Digitalis purpuread)是一个很好的例子毛地黄的M等位基因控制合成红色花花色素苷。D为修饰基因,它控制合成大量的花色素苷,花呈深红;d只能控制合成少量的花色素苷,花呈浅红。W座位控制花色素在花冠上的不同分布。W等位基因使红色色素只分 ...
抑制基因是一种非常重要的互作。此是一个基因抑制另外一个基因的表达,但其生化途经和显性上位不同。在报春花属(Primula)中K基因可以控制合成一种黄色的锦葵色素,但另一个D基因存在时可抑制其表达。 ...
在减数分裂时同源染色体之间的交换导致了遗传重组的发生。一个交叉就是交换的位点。交换是同源染色体通过联会、断裂、重接在相关位置彼此交换。遗传重组的证据是在减数分裂中发现交换时细胞学标记发生交换,遗传标记也发生交换交换是一个交互事件,在真核中于减数分裂的前期 I 可发生在二价体的阶段。重组是交换的结果,但交换并不一定引起重组,如两标记间发生偶次数交换,标记并不发生重组。所以一般实际的交换值要大于重组值 ...
1910年摩尔根和他的学生布里吉斯(Bridges C.B)研究了两对基因的遗传,发现了连锁和互换,建立了遗传学第三个基本定律,连锁法则。 摩尔根发现果蝇的红眼和紫眼(purple)、长翅和残翅(vestigial)两对性状都是非伴性性状,各自的遗传都符合孟德尔法则。他们将红眼长翅的果蝇和紫眼残翅的果蝇进行杂交,F1代为红眼长翅,然后将F1和双隐性亲本进行测交,所得到的测交后代按孟德尔法则应有四种 ...
染色体内重组是交换的结果,但交换不一定导致重组。因为我们是依据标记基因来判断是否重组的,若在两个标记基因之间发生奇次数交换,结果必然导致重组,但若发生偶次数交换,标记基因并不重组。不能进行自由组合的基因群则称之为连锁群(Linkage group),排列在同一条染色体上及其同源染色体上的基因同属于同一连锁群。一种二倍体生物的连锁群数应和其染色体对数相等。在少数的生物中如雄果蝇和雌家蚕都不发生重组, ...
摩尔根的果蝇实验已表明重组是同源染色体发生交换的结果,两个连锁基因之间的距离决定了它们的重组值。那么我们能否用实验的方法来确定真核生物染色体上不同基因的位置呢,答案是肯定的,这种方法我们称之为遗传作图。
基因位于染色体上,而且位置相近的基因是相互连锁的,不难想象基因在染色体上是直线排列的,但如何来证明这个问题呢?摩尔根的学生斯特蒂文特(Sturtevant)想出了一个巧妙的办法,那就是三点测交(three-point test cross)。从几何知识上知道,要证明a、b、c三点共线,可以通过三点之间的距离加以证明,当ab bc=ac时三点在一条直线上。斯特蒂文特借鉴了这一思路,选择了六个性连锁基 ...
如果在二条非姊妹染色单体上分别发生两次单交换,单交换Ⅰ的概率为α,单交换Ⅱ的概率为β,如果这两次交换同时发生(即为双交换),其概率理论上为(α)(β),假设实际测得双交换的概率为γ。并发系数(coefficidence of coincidence or coincidence)C为观察到的双交换数除以预期双交换这一比值。 按上列数据C=γ∕(α)(β)这个结果远小于1,也就是说实际发 ...
在减数分裂的前期,二价体是有4条染色单体,交换并不限于二条非姊妹染色单体之间,而有各种不同的交换形式。为了便于说明,我们巴4条染色单体分别标上序号。当2-3之间已发生了一次交换的基础上,再发生2-3二线双交换;1-4四线双交换;1-3三线双交换和2-4三线双交换机会是均等的。一般说来第一次交换仅对于2-3二线双交换有干涉,对其他类型的双交换没有干涉。 ...
链孢霉的野生型又称为原养型(prototroph),在基本培养基上就能生长,子囊孢子按时成熟呈黑色。有一类突变型是不能合成某些物质,称为营养缺陷型(auxotroph),在基本培养基上不能生长。如赖氨酸缺陷型是自己不能合成赖氨酸,因此培养时必须在基本培养基上加入适量的赖氨酸,此种缺陷型才能生长。缺陷型比野生型的子囊孢子成熟得慢,所以镜检时突变型的子囊孢子呈白色,和黑色的野生型子囊孢子很容易分辨。 ...
表示核酸长度的单位。1000碱基(对)的长度为1kb。为kilobace的缩写。 ...
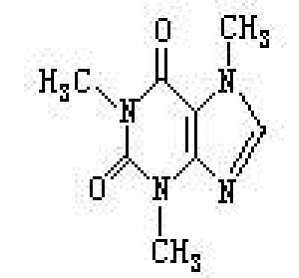
C8H10O2N4·H2O,是一种生物碱,亦称茶素或茶碱(theine)等。熔点238℃,在178℃升华。溶于水呈苦味。呈现对嘌呤衍生物共同的红紫酸铵反应。存在于咖啡的种子、柯拉树(Kola)的果实、可可的种子和茶叶中。具有对中枢神经系统的兴奋作用,以及利尿等作用。除了对肾的作用、对心肌的兴奋作用以外,也有对平滑肌特别是对支气管、冠状动脉的抑制(舒张)作用。据认为抑制水解cAMP的环化磷酸二酯 ...
亦称3,4-二羟基桂皮酸。在咖啡豆中,虽与奎尼酸(quinic acid)形成结合物(氯原酸)大量存在,但在一般高等植物中常以游离酸、甲酯、奎宁酸,少数情况下以葡萄糖结合物的形态广泛存在。易受酚氧化酶的氧化,是组织发生褐色变化的重要原因之一。对植物的生长有抑制作用,是生长抑制物质——抑制剂β的成分之一。可抑制过氧化物酶、吲哚乙酸氧化酶的作用。在生物体内,它是由L-苯丙氨酸脱氨基后生成的反式桂皮酸逐 ...
亦称k粒。它存在于放毒型草履虫的细胞质中,能进行自体复制,显有与病毒相似的性质,一般认为是寄生性病毒。卡巴粒的直径为0.2—0.5微米,长度为0.5—5微米。在具有放毒作用强的放毒型中,一个细胞内有400—1600个卡巴粒,这种粒子也可向草履虫的培养液中释放,可杀死摄取这种粒子的敏感型草履虫。与卡巴粒相类似的粒子有λ、σ、γ、δ等粒子。 ...
将实验获得的数据与理论估算值进行对比,检验两者符合程度的一种测验方法。例如在遗传实验测验时,实验分离比率与理论比是否一致,调查两种性状之间有无关系时,就往往使用这种方法。χ平方一般可按下式求算:χ2=∑(观察数-理论数)2/理论数。下表是豌豆杂交试验(黄粒×绿粒)F2的分离比测验的一个例子: 从χ2表上,查出自由度所关系(比较项数减一)的数值,此数即为该偏差出于偶然机会的概率。上例中,自由度 ...
即Bacille de Calmette Guérin的缩写。是由法国的卡尔米蒂(A.Calmette)和介林(C.Guér-in)从1908年起13年间将牛型结核菌,以牛胆汁马铃薯为培养基,累代培养得来的弱毒性结核菌。广泛应用于预防结核病发作。因为对于结核的死菌免疫无效,所以用此弱毒性活菌接种于人体(所谓BCG接种),对结核予以人为的免疫。接种BCG会在局部及所属淋巴结等处出现结核性变化,但其反 ...
系由卡那霉素链霉菌(Streptomyces kanamy-ceticus)。产生的类似于链霉素的抗菌素,是梅泽浜夫等(1957)分离得到的。对结核菌及其它细菌具有抗菌作用。结合于细菌的30S的核糖体,可引起mRNA的错译,在蛋白质生物合成的初始阶段起到抑制作用。 ...
关于丁香通
公司信息
个人用户
企业机构


