


指有形象视觉的眼分辨空间物点位置的能力,是视空间(参见空间觉)形成的基础。虽然可区分为两种不同的视力:方向视力(辨别物空间方向,即视野上的位置的差异)和深度视力(辨别离眼的距离的大小),但如果单说视力,通常是指前者。这无非就是眼作为光学仪器的分辨能力。能够辨认的最小点的视角(绝对阈值)或辨别间隔最近的两点的视角(辨别阈值)倒数表示,以视角1′定为视力1。正常人的视力(按辨别阈值)为1—2。视力 ...
这种学说认为脊椎动物的视细胞中锥细胞与昼间视觉有关,主司色觉;另一方面,杆细胞主司黄昏视觉、无色觉。经舒尔茨(M.Schuetze,1866)的解剖学研究、帕里纳德(H.Parinaud,1881)的病理学研究、克里斯(V.Kries,1895)的生理学研究,至今已成定论。在网膜中央凹唯有锥细胞而无杆细胞,越向周边区锥细胞越少,杆细胞渐多,与这种解剖学观察相适应,肯定二元说者尚有许多生理学、心 ...
以视黄醛为载色基因的视色素的吸收峰虽然在不同动物和杆细胞、锥细胞中都有所不同,但对于波数(波长的倒数),被标示的吸收曲线主吸收带的形状却是完全相同的。达特纳尔(H.J.A.Dartnall,1953)用计算图表表示了吸收峰与各波长吸收率的相关关系。如果使用这种图表,即使抽提的视色素不纯,只要知道吸收峰的波长就可以求得吸收曲线。对视黄醛2系的视色素也已按同样的原则制作了计算图表。 ...
指大脑皮层的初级视觉中枢(参见感觉区)。从外侧膝状体发出的视觉二级神经元形成视放射,终止于大脑枕叶的所谓纹区〔striate cortex,布罗德曼(Brodmann)脑定位图第17区〕,这一部分,称为大脑皮层视区。因为在各部位神经纤维均排列得极有规律,所以来自左右网膜相应点的视神经纤维,经各自的神经通路都汇聚于视区的同一神经元。相当于网膜黄斑区的皮层视区的面积明显地扩大,其比例可达一万倍,越 ...
传递视觉的神经,在脊椎动物,一直被认为是第二脑神经,可是原来曾将之与网膜一起作为脑的一部分。因此也被称为视束。网膜的第三神经元集合于视乳头,贯穿巩膜的筛板,左右各成一条神经束。为连接视神经脑膜的视神经鞘所包裹,穿过视神经管进入颅腔。在间脑漏斗部的前面,进行所谓视神经交叉后,称为视索(tractus opticus)。在头索类的前脑,仅有眼点,没有视神经。鱼类的视神经交叉常常是在脑内进行。在低等 ...
亦称视束交叉。即指把进入头骨的视神经(视束)在接近间脑前方(鱼类在脑内)的交叉。在低等脊椎动物中,交叉后成为视索的左右视束全部走向各自对侧的中脑;在哺乳类,左右视神经的内侧一半交叉,外侧一半为同侧视索达至间脑的后侧膝状体(半交叉)。因此,从印在左右网膜上的同一象的成象部位所发出的视神经,可以到达脑内同一部位,这是与两眼的网膜象成为单一象的认识有关。 ...
包含于网膜视细胞外段的感光蛋白。由视蛋白(蛋白质部分)和11-顺视黄醛(载色基团)组成,视色素吸收光即产生光感觉。动物具有与光环境相适应的固有的视色素,这些视色素可按其吸收特性(颜色)来分类。陆生和浅海动物一般具有吸收峰在500毫微米左右的紫红色视色素(视紫红),而深海动物具有吸收峰在短波侧约480毫微米的金黄色视色素(视金,crysopsin),淡水鱼类、两栖类的视色素吸收峰在520毫微米以 ...
指复眼视细胞轴突在视叶第一神经节的神经节层(lamina ganglionaris)二次神经与突触结合的部位。因大致是与单眼对应所形成的新的构造单位和功能单位,所以也称为神经单眼(neurommatidi-um)。例如具有感杆束分离型复眼的蝇类的视神经束,与来自相邻的每条由6个细胞组成的共计6条视细胞轴突一起进行二次神经与突触结合。 ...
指神经细胞及神经纤维集合而成的结节状构造。在脊椎动物,存在于网膜的最内层(光进入的方向),而在软体动物的眼里,则在网膜的最外层。但在甲壳类及昆虫类的复眼中,是从个眼的视细胞层分开成为另外的构造,一般具有视神经层(laminaganglionaris)、外髓(medulla externa)、内髓(medulla interna),在甲壳类还接有终髓(me-dulla terminalis),所 ...
为两栖类、鱼类和圆口类的间脑视丘下部视神经交叉的前方至上方存在的神经分泌细胞细胞体之集合体。其神经分泌物质可被Gomori染色法的铬矾-洋苏木素或三聚乙醛-复红染色。含神经性脑下垂体激素。即脑下垂体的神经叶激素在此处合成,由轴突运至神经叶。在爬行类以上的脊椎动物此核二分,分别称为视束上核和室旁核。 ...
脊椎动物哺乳类、鸟类和爬行类丘脑下部(间脑的底部)的神经核(神经细胞细胞体集中的部分)之一,位于视神经交叉的上方。由神经分泌细胞所成。其神经分泌物质对Gomori染色法的铬矾-洋苏木素及三聚乙醛-复红为阳性。含神经性脑下垂体激素。神经叶激素在此处合成,从其轴突运至脑下垂体神经叶。其近旁的室旁核也具有同样的功能,但两栖类、鱼类和圆口类区别不出此二核,只有一个视束前核。在哺乳类动物含催产素的细胞与 ...
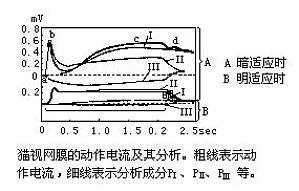
亦称视网膜动作电位图。网膜受光照时产生电位,把这种电位记录下来便是视网膜电位图,简称ERG。勿从杜波依斯-雷蒙(E.H.Dubois-Reymo-nd,1849)发现网膜静止电流和霍姆格伦(F.Holm-grem,1866)发现动作电流以来,为了探讨网膜的视觉机制把ERG作为有用的客观指标,对哺乳类、两栖类、爬行类、鱼类、头足类等动物进行了广泛的研究。把网膜放在黑暗中发现,产生的静止电位的极性 ...
头足类的感光色素蛋白质,是原富之夫妻(1965)在鱿鱼的视网膜中发现的。吸收曲线和视紫红质相似,但以全反式-视黄醛为发色团,吸收光后11-反式视黄醛游离。被认为是供给对视物质再生所需要的11-顺式视黄醛的光异构酶。视物质存在于感杆小体(rhabdomere)上,而这种色素多存在于视细胞的内节。 ...
指在动物的感光细胞中分化出有特殊视觉机能的细胞,即所谓光感受器细胞。这些细胞有的以所谓分散光感觉器的形式,分散在身体表面,有的由很多的细胞集合在一起构成网膜,参与眼的组成。后者被称为视细胞或视网膜细胞。视细胞以初级感觉细胞的形式出现,借助视细胞中的感光色素的光化学反应产生的兴奋,传送至传入性神经纤维(视神经)。通常,将此细胞看作是能量转换的场所具有特殊的形态分化。尤如蚯蚓的分散光感觉器官,有的 ...
指随着明暗的适应状态,脊椎动物视网膜的视细胞(特别是锥细胞)和色素上皮细胞的色素(暗褐菌素,fuscin)出现移动的现象。在摘出眼和剥离的视网膜也可观察到这种现象。这是眼的一种物理性适应,有时是与固有的适应(生理性适应)过程相对应而言的。在暗适应时,杆细胞位于外境界膜,色素位于色素上皮细胞的底部,而锥细胞则位于杆细胞和色素中间的位置,此称为暗位(德Dunkelstellung)。在明适应时,锥 ...
在人体生理学上,在眼球不动的状态下,盯准眼前的一个目标,此时固视(fixation)所及的范围称视野。也就是投影于网膜感受面的视空间(visualspace)。如果注视某一物体点,通过眼球的适当活动,在网膜看得最清楚的中心部位,即中央凹形成其鲜明的物象。这称为中央视觉或直接视觉。同时,该点四周的物象,即离开视野中心的部位(边缘部),成象较不鲜明,而称为边缘视觉或间接视觉。中心部是明适应感光和视 ...
为脊椎动物视网膜的细胞。视锥细胞的外端(背光端)呈圆锥状突,作为该细胞的感光性部分,可与视杆细胞突相比;在昼行性动物的视网膜上视锥细胞特别多,因此被看作是具有白昼视觉、色觉的视细胞因素(昼间视觉装置)。在低等脊椎动物的眼中,视锥细胞的柄部在白昼光下,可将收缩的视锥细胞突引到视网膜的成像面。视锥细胞的电子显微镜下的内部构造与视杆细胞相似,视锥细胞的锥盘(cone di-sk)与长轴呈直角方向重叠 ...
亦称视线运动反应或(运)动视反应。当被视物体或视野移动时,为使视网膜象尽可能保持稳定不动所进行的眼球、头部乃至整个身体的运动。在脊椎动物、昆虫、高等甲壳类、头足类,均可见到这种反应。此时的视运动反应的控制系统是以视觉目标为输入,视线为输出的随动系统(servo)机制,构成以视标与视线的差异为误差信号而活动的反馈控制系统。一般来说,视运动反应对于追踪运动物体和对抗流动的媒介体而保持原位行为起着中 ...
脊椎动物视网膜的视细胞、杆细胞外段等所含的视色素,是以11-顺视黄醛为生色基因,分子量约40,000的色蛋白。蛙、牛的视杆细胞外段,它占构成蛋白质的80%以上。把视网膜磨碎,悬浮于36%蔗糖液中,远心分离,收集上浮的杆细胞外段,可在表面活性剂溶液中抽提视紫红。视觉的初始过程,是由视色素的光反应而引起外段膜的Na 离子的透性被受抑制。视紫红的光反应过程是通过低温抑制热反应,对随温度逐步上升而出现 ...
氨是植物的重要的氮化物的来源,但同时也是生物体的一种毒性物质。从土壤中吸收氨,在土壤pH值较低时尤为显著,容易造成氨的积累;植物一般容易呈现中毒;如果碳水化合物不足,而这种症状更为严重。然而在细胞液中含有多量有机酸的植物,例如酢浆草、酸模、秋海棠等,由于细胞液中酸度较高,所以即使氨被过量的吸收和积累,也会因全部成为铵盐而被解毒,故不会引致中毒,这种植物称为适酸植物或适氨植物。 ...
关于丁香通
公司信息
个人用户
企业机构


