


网络 第二节 免疫球蛋白基因的结构和多样性 表3-2 免疫球蛋白基因定位 编码多肽链 基因符号 (人) 基因定位(染色体) 人 小鼠 κ轻链 IGK 2p11 6 λ轻链 IGL 22q11 16 ...
网络 三、CD、粘附分子与Ig超家族的关系 免疫分子的命名根据不同的角度往往有不同的归类和命名,图3-3归纳了人白细胞分化抗原(CD)、粘附分子(AM)、免疫球蛋白超家族(IGSF)、细胞因子受体(CKR)、补体受体(CR)以及主要组织相容性复合体抗原(MHC)等免疫分子命名的相互关系。 (1)A表示CD命名范围中的粘附分子,如CD49a ...
网络 二、Ig超家族的特点 (一)Ig超家族的结构特点 Ig超家族成员均含有1~7个Ig样结构域,每个Ig样结构域约含70~110个氨基酸残基。其二级结构是两个各含3~5个反平行β折叠股所形成的β片层(anti-parallel β-pleated sheet)平面,每个反平行β折叠股由5~10个氨基酸残基组成,β片层内侧的疏水性氨基酸起 ...
网络 第三章 免疫球蛋白超家族 免疫球蛋白基因的研究近年来获得重大突破。日本学者利根川进(Tonegawa)因在免疫球蛋白基因结构研究有突出贡献而获得1987年诺贝尔医学和生理学奖。应用X结晶衍射分析、DNA序列分析和园双色等技术研究表明,许多细胞膜表面分子和机体某些蛋白分子多肽折叠方式与Lg相似,在氨基酸组成上与免疫球蛋白可变区(V区)或/和恒定区( ...
网络 二、C4结合蛋白 C4结合蛋白(C4bp)是一种含量丰富的可溶性血清糖蛋白,分子量为550kDa,1977年由Ferreira等所报道。其分子结构模式现多以Dahlback等(1983)描述的“蜘蛛样”(spiderlike)结构来分析其结构及功能。C4bp由8个亚单位组成,电镜下观察形似蜘蛛,其中有7条分子量相同(均为70kDa)的长链(α ...
网络 第二节 补体调节分子的结构及功能 补体系统的激活为一种级联反应,但受到多种调节分子的严格控制,其反应的程度和单一成分的反应都是在生物反馈近代制下而进行的,从而限制了活化的扩大化,以维持补体水平的平衡。调节作用包括两个方面,即自身衷变失活及一些抑制物的灭活作用。前者指已活化的补体分子均不稳定,如不及时与靶细胞膜结合即迅速衰变失活;后者是通过抑制物的 ...
网络 十一、P因子 P因子又称备解素(properdin),是替代途径中除C3以外最先发现的一种血浆蛋白。现已探明,P因子以聚合体形式而存在:即三聚体(54%)、二聚体(26%)和四聚体(20%)都有,但特异活性的顺序依次为:四聚体三聚体二聚体。P因子为由4条相同的肽链(分子量各55kDa)组成的四聚体分子,链间以非共价键相连接,分子量为220kD ...
网络 十 、D因子 D因子是启动替代途径激活的重要成分,为由222个氨基酸残基组成的单链丝氨酸蛋白酶,分子量仅25kDa。D因子在血清中的浓度很低(1-2μg/ml),主要以活化形式而存在。但可能还有一种以酶原形式而存在的由239个氨基酸残基组成的D因子。具有活性的D因子(D)可能在第234-235位的精氨酸-赖氨酸键处将B因子裂解为Ba和Bb两个 ...
网络 九、B因子 B因子(factor BBf)替代激活途径中的重要成分,由Blum于1959年首先发现。B因子为由733个氨基酸残基组成的单链糖蛋白(糖含量约7%),分子量93kDa。由于这些氨基酸的迂回折叠形成三个大小相近似的球形区。其中1个为Ba,其余两个呈哑铃状为Bb。Bb中靠近N端的一个球形区可同C3b结合,另一个球形区可能是催化区(图5 ...
网络 八、C9分子 C9是形成膜攻击复合体(MAC)的最后个分子,为一单链糖蛋白,分子量79kDa。经对cDNA推导的氨基酸序列分析发现,C9为一两性分子。C端37kDa由疏水性氨基酸组成称C9b,N端34kDa由亲水性氨基酸组成称C9a因此C9以其羧基端部分嵌入细胞膜的脂质双层中。而N端则为与c 5b-8相结合的结构域。C9具有自发聚合的作用,但 ...
网络 七、C8分子 C8是由α、β、γ三条肽链组成的三聚体糖蛋白,分子量为155kDa。其中α链和β链均为64kDaγ链为22kDa。α链和γ链间以二硫键共价结合,而α链与β链间则为非共价键结合(图5-10)。C8分子中也含有TSP-1和LDL受体结构功能域。在C8α和C8β多肽链的中央(157-501个氨基酸残基间),几科不含半胱氨酸残基,为与细 ...
网络 六、C6和C7 C6和C7有许多相似之处,均为单链糖蛋白,且分子量也相近分别为128kDa和121kDa。编码C6和C7分子的基因可能由共同的祖基因进化而来。C6和C7在氨基酸水平上有33.5%的同源性。近几年来,对C6的结构及功能进行了较深入的研究,由cDNA序列推导成熟C6的全部多肽链含有913个氨基残基,前面还有21个独特氨基酸残基组成 ...
网络 五、C5分子 C5是形成膜攻击复合体(MAC)的第1个补体分子。C5由以二硫键相连接的α、β链组成,分子量190kDa,其中α链为115kDa,β链为75kDa(图5-9)。C5与C3和C4的结构相类似,但没有链内硫酯键。靠近N端的第74-75位精氨酸一亮氨酸键为C5转化酶作用的部位。在C5转化酶的作用下,C5α链N末端裂解出一个分子量为1 ...
网络 四、C3分子 C3处于两条激活途径的汇合点,在补体系统活化过程中起着枢纽作用,并为替代途径激活的关键分子。C3的α、β两条肽链组成,之间以二硫键相连结,分子量为195kDa,其中α链为115kDa,β链为75kDa(图5-6)。其在血清中的含量高于其它补体分子,约为0.55-1.2mg/ml。同C4分子一样,C3分子的α链在半胱氨酸和谷氨酸残 ...
网络 三、C2分子 C2的序号似是补体的第2个成分,但在经典激活途径的激活顺序上却在C4以后被活化。C2分子的一级结构已全部搞清楚,它是由723个氨基酸残基组成的单肽链糖蛋白,分子量约110kDa(图5-5)。当C2与已固定于细胞膜固相上的C4b结合为复合物时,C1s丝氨酸蛋白酶可从C2肽链的精氨酸和赖氨酸(223-234)间,将C2裂解为两个片段 ...
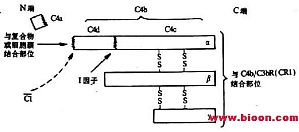
网络 二、C4分子 C4是经典激活途径中第二个被活化的补体成分,分子量约为210kDa,由α(90kDa)、β(78kDa)及γ(33kDa)三条肽链借二硫键连接组成(图5-4)C4的分子结构较为特殊,其α链中含有一个在半胱氨酸和谷氨酸残其间形成的内硫酯键。α链的N端有C1s丝氨酸蛋白酶的作用点。当C1s将C4α链的精氨酸-丙氨酸键(76-77位) ...
网络 第五章 补体的分子生物学 补体系统由30多种蛋白分子所组成,是迄今所知机体中最复杂的一个限制性蛋白水解系统(limited proteolysis system)根据各成分功能不同,将它们分为三组。第一组为补体系统的固有成分共14个蛋白分子。即C1(含三个亚组分:C1q、Clr和Cls)、C4、C2、C3、C5、C6、C7、C8、C9、B因子、D ...
网络 四、 核酸标记技术检测法 通过核酸标记技术可将细胞因子cDNA作为基因探钊检测细胞内细胞因子基因组DNA或mRNA。主要有以下几种方法: 1.应用同位素(或非同位素)标记的cDNA探针,检测经Northern blot后细胞因子mRNA水平或采用打点杂交法。 2.应用标记cDNA探钊与细胞或组织切片进行原位杂交,然后进行放 ...
网络 三、胞浆或胞膜细胞因子(或受体)的检测 许多细胞因子产生过程中,有一个胞浆到胞膜,再释放到体液或培养上清中的动态变化。一般可采用免疫组织化学染色技术或免疫荧光技术检测细胞或组织切片中细胞因子(或受体)定位(胞浆、胞膜),产生细胞的频数,以及胞浆、胞膜、上清中浓度改变的动态变化。目前已报道IL-2、TNF-α、IFN-γ等细胞因子以及几科所有细 ...
网络 二、免疫学检测法 免疫学检测法的基本原理是细胞因子(或受体)与相应的特异性抗体(单克隆抗体或多克隆抗体)结合,通过同位素、荧光或酶等标记技术加以放大和显示,从而定性或定量显示细胞因子(或受体)的水平。这类方法的优点是实验周期短,少受抑制物或相似生物池功能因子的干扰,如抗体的特异性高可区分不同型或亚型的细胞因子(如IFN),一次能检测大量标本, ...
关于丁香通
公司信息
个人用户
企业机构


