

- ¥100 - 1000
- Xybio
- 上海
- P1856
- 2026年05月27日

企业认证
相关产品推荐更多 >

万千商家帮你免费找货
0 人在求购买到急需产品
相关阅读
对话丨深圳先进技术研究院赵乔团队的「可编程植物」探索abinScience 体内级抗体,年度锁价,低至 1200五大应用案例:Mustang Q 膜层析应用全解析1 个小工具,一次性搞定流程图、质粒图谱和信号通路图- 详细信息
- 文献和实验
- 技术资料
- 保存条件:
-20
- 保质期:
2年
- 英文名:
pSwitch
- 库存:
100
- 供应商:
上海烜雅生物科技有限公司
- 规格:
干粉/液体
名称:pSwitch果蝇表达质粒
别称: pSwitch
| 原核抗性: | Amp |
|---|---|
| 筛选标记: | Hyg |
| 克隆菌株: | DH5a |
| 培养条件: | 37度 |
质粒属性
| 载体宿主: | 昆虫细胞 |
|---|---|
| 载体用途: | 蛋白表达 |
| 基因种属: | |
| 基因类型: | ORF |
| 原核抗性: | Amp |
| 筛选标记: | Hyg |
| 荧光蛋白: |
质粒简介
pSwitch质粒由上海烜雅生物科技有限公司提供
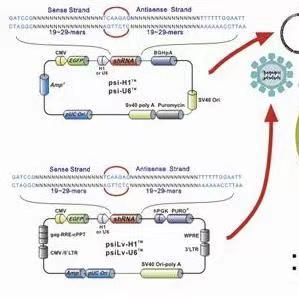
质粒图谱
质粒序列:详询
质粒菌株产品操作说明书
一、扩增流程
收到产品后,请先根据产品管壁标签来判断产品形式,并在扩增前准确查找该质粒菌株的抗性、感受态和培养温度。
1、质粒干粉(常温运输,存于-20度,90天保质期,请务必转化挑单克隆培养,不要直接使用和测序)
①收到质粒干粉后请先5000rpm离心1min,再加入20μl ddH2O去离子水溶解质粒;
②取1支100μl 感受态于冰上解冻10min,加入2μl质粒,再冰浴30min后,42℃热激60s,不要搅动,再冰浴2min;(从第二步开始均要在超净工作台中无菌操作)
③加入900μl无抗的LB液体培养基,180rpm震荡37℃培养45min;
④6000rpm离心5min,仅留100μl上清液重悬细菌沉淀,并涂布至目标质粒抗性的LB平板上;(可使用本平台的平板涂布专用玻璃珠进行涂布,可以比传统涂布方法获得更多转化子)
⑤将平板正向培养1h,再倒置37℃培养14h。如果要求是30度则培养20h;
(菌落过多则将质粒稀释后再转化。没有菌落则加入10μl质粒转化。另不要直接转表达感受态,要先转克隆感受态,重提质粒后再导入表达感受态)
⑥挑取单菌落至LB液体培养基中,加入对应抗生素,220rpm震荡培养14h,根据实验需要和质粒提取试剂盒说明书提取质粒。
2、甘油菌种(冰袋运输,存于-80℃,保质期90天,请务必划线挑单克隆培养)
四区划线后挑单菌落培养,酵母菌需要先液体复苏再四区划线,再挑单菌落液体培养。
3、穿刺菌种(冰袋运输,存于4℃,保质期7天)
穿刺接种,液体培养后四区划线,再挑单菌落液体培养。
4、菌落平板(冰袋运输,存于4℃,保质期7天)
直接挑取单菌落至液体培养基中。
5、液体质粒(冰袋运输,存于-20℃,保质期90天)
单独提取的液体质粒收到后可直接使用。
6、滤纸质粒(常温运输,存于-20度,90天保质期,请务必转化挑单克隆培养,不要直接使用和测序)
收到货后将滤纸画圈部分剪下放入EP管中,加100ul无菌水将滤纸浸湿并浸泡5min,吸取5ul质粒转化,离心全涂。
二、转化图片

| P4037/pCMV-SPORT6-PELI1人源基因质粒 |
| P4038/pCMV-SPORT6-IFITM2人源基因质粒 |
| P4039/pCMV-SPORT6-MAFF人源基因质粒 |
| P4040/pCMV-SPORT6-ARMCX2(1同义突变)人源基因质粒 |
| P4041/pCMV-SPORT6-TMEM183A人源基因质粒 |
| P4042/pCMV-SPORT6-LRIF1(603-2310bp)人源基因质粒 |
| P4043/pCMV-SPORT6-GRAMD2B人源基因质粒 |
| P4044/pCMV-SPORT6-OXCT1人源基因质粒 |
| P4045/pCMV-SPORT6-UTP15(1同义突变)人源基因质粒 |
| P4046/pCMV-SPORT6-RPS3A人源基因质粒 |
| P4047/pCMV-SPORT6-RAB38(1同义突变)人源基因质粒 |
风险提示:丁香通仅作为第三方平台,为商家信息发布提供平台空间。用户咨询产品时请注意保护个人信息及财产安全,合理判断,谨慎选购商品,商家和用户对交易行为负责。对于医疗器械类产品,请先查证核实企业经营资质和医疗器械产品注册证情况。
 文献和实验
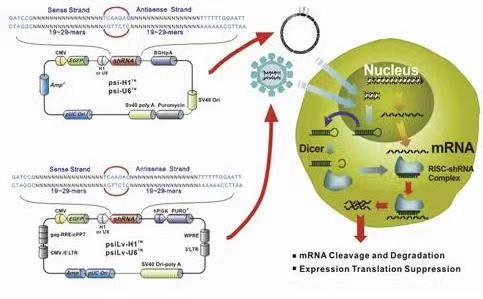
文献和实验有弊。制备某个家族缺陷的转基因小鼠变得相当容易。但如果你想要了解某个miRNA的特定功能,那就得将与之接近的miRNA全部去除。 miRNA海绵的应用潜力无穷。它可用于靶点预测的验证,也能分析miRNA的功能丧失表型。由于miRNA海绵是由表达质粒编码的,那么它也很容易推广到其它形式,譬如慢病毒、腺病毒或转基因,适合各种不同的细胞类型。Sharp还在研究中证实,在UTR上多加几个miRNA结合位点,能增加反义序列的量,从而提高海绵的效力。从原理上来说,miRNA海绵的CMV启动子也可以换成其它组织
使用也能扩增RNAi的基因沉默效应。 科研工作者已开始RNAi效应增强子和效应抑制子的研究,认为RNAi是机体细胞一种强有力的基因调控机制。但是在哺乳细胞中外源导入大分子dsRNA(>30nt)常常导致非特异性的基因沉默效应,因为它激活了核糖核酸酶RNaseL而导致非特异性的RNA降解。相反,siRNA( 目前外源注射和转染是转运siRNA到细胞或机体的主要途经,之后基因沉默效应可以持续数天,同时传至下一代细胞,但终将短时间内消失,因此,最近许多科研小组开发了siRNA的表达质粒,通过瞬时
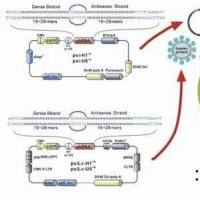
双链短RNA(dsRNA)阻断基因表达机理及双链RNA的构建
相关专题 1998年以前,研究者们都认为正义RNA和反义RNA 都可以阻断基因的表达,直到Andrew Fire发现双链RNA才是阻断基因表达 的因素,这种阻断作用称之为RNA,它被评为2001十大科学成就之一。 随后的研究中发现,RNAi现象广泛存在于线虫,果蝇,斑马鱼,真菌以及植物等生物体内,这些生物体利用RNAi 来抵御病毒的感染,阻断转座子的作用。RNAi能高效特异的阻断基因的表达,在线虫,果蝇体内,RNAi


