
质谱成像 用一项新兴的非常有效的质谱技术结束本章看起来是很合适的,这项技术就是质谱成像(imaging mass spectrometry,IMS)。在这个最新的进展中,基质辅助激光解吸电离飞行时间质谱(MALDI-TOF-MS)对直接来源于组织切片中的肽和蛋白质进行剖析和成像,以便获得有关这些肽和蛋白质局部相对丰度和空间分布的精确信息。这种成像实验的结果能够提供大量的信息,利于研究者测量 ...
蛋白质组分析方法--MS/MS 由于当前的质谱仪器能够特异性地选择和拆碎肽,在计算机的帮助下,可以利用源后衰变(post-source decay,PSD)或碰撞诱导解离(collisioninduced dissociation,CID)获得的图谱来测定序列标签和肽的全序列。另外的肽序列信息使蛋白质鉴定减少了不确定性,而且在蛋白质没有列入蛋白质序列数据库时,可以搜索表达序列标签(expr ...
蛋白质鉴定工具 蛋白质鉴定意味着将实验测定的蛋白质性质和可以利用计算机手段从一个数据库得到的蛋白质性质进行比较。考虑到这些不同的性质,已经开发了一大批蛋白质鉴定工具,如氨基酸组成、序列标签或质谱衍生的数据等,所有的这些工具都是通过与蛋白质数据库中的某一个蛋白质条目提供最佳的匹配来达到鉴定蛋白质的目的。 以PMF和MS/MS为基础的蛋白质鉴定工具只能对部分实验信号进行注释或解释。当前已 ...
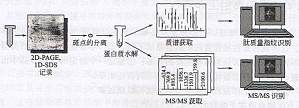
蛋白质组学经典的基子凝胶的方法 从历史观点上来说,蛋白质组学与利用二维凝胶电泳技术的大规模蛋白质鉴定和质谱紧密相关,其中电泳作为分离方法,质谱作为后续的分析工具。尽管这种方法已经不能涵盖在复杂的生物样品中发现的所有蛋白质,但是它依然在相当多的实验室中使用。在这种处理方法中,蛋白质样品应遵从于二维聚丙烯酰胺凝胶电泳(2D―PAGE)的高分辨率和分离能力。二维凝胶电泳是由O’Farrel(19 ...
蛋白质组学分子扫描仪方法 经典方法的特征是用高分辨率的技术分离蛋白质,然后在几个月内这些蛋白质都可用于分析。这种经典方法的一个瓶颈是与繁重的样品处理有关,即在2D-PAGE上对大样品进行所有组分的系统鉴定。在分子扫描仪方法(Binz等,1999a;Bienvenut等,1999)中,通过1D―SDS或2D―PAGE分离的蛋白质同时消化,并通过电转移到一个收集膜中。在膜上,消化的肽浓缩成一些 ...
蛋白质组学表面增强激光解吸电离方法 表面增强激光解吸电离(surface-enhanced laser desorption/ionization,SELDl)技术基于MALDI技术,利用了其测量部分。SELDI涉及一个在特殊化学表面的亲和俘获过程,随后利用激光解吸电离探测进行质量分析(Merchant和Weinberger,2000)。 SELDI过程可以描述为4个部分。 ...
蛋白质组学同位素标记的亲和标签方法 Aebersold和其合作者开发了一种同时对蛋白质组进行鉴定和相对定量的方法,即一种包含在无胶蛋白质组流程中的同位素标记技术(Gygi等,1999)。他们提出使用一种试剂,这种试剂能够使两种同位素在形式上具有明显的区别,即同位素标记的亲和标签(isotopicallycodedaffinitytag,ICAT)试剂。ICAT试剂的结构包含一个生物素头部、 ...
蛋白质组学多维蛋白质鉴定技术 面对1D―SDS和2D―PAGE技术在回收胶中质量极端(很大或很小)、疏水性很强或pH值很高的蛋白质时遭遇到的某些困难,Yates和其合作者(Wolters,Washburn和Yates,2001)发展了一种非基于凝胶的方法,即采用一种所谓的鸟枪法技术来鉴定尽可能多的蛋白质。多维蛋白质鉴定技术(multi-demensional protein identif ...
基因克隆将基因组信息翻译成某种蛋白质组学信息 将基因组信息翻译成某种蛋白质组学有用信息的第一个界面是在表达的克隆阶段。通过普通数据库可以获得大量的有关基因组的信息。挖掘这些信息可以用来预测开放阅读框(open reading frame,ORF),这些ORF可以根据序列的相似性预测功能而后归类。但是为了验证这些假定蛋白,必须先完成一项困难的任务,即获得一个预测ORF的真实克隆,并将其转化为 ...
表达蛋白 在获得为表达准备的克隆之后,可以利用96孔板小规模地表达这些目的蛋白来选择一些能生产可溶蛋白质的克隆。传统方法(Doyle等,2002)和商用的机器人自动仪(Lesley,2001)技术都可以实现高通量表达和纯化毫克级的蛋白质,编者预期的大规模处理方法是发展一种特别设计的自动化方式。表达毫克级的蛋白质利用传统方法完成至少需要发酵500~1000ml的培养液,如果将96个蛋白质同时 ...
高通量蛋白纯化 为了尽可能地提高蛋白质的纯度,而同时又避免纯化过程中过大的工作 强度,习惯于在目标蛋白质上贴上一个纯化标签。Stevens(2000)的文章给出了一个全面的可用融合标签的目录。一般来说,比较长的标签,如硫氧 还蛋白(La Vallie等,1993)、麦芽糖结合蛋白(山Guan,Riggs和Inouye,1988)和谷胱甘肽转移酶(Smith和Johnson,1988 ...
蛋白表达纯化流程的确认和展望 为了检验表达系统和纯化流程的鲁棒性,研究人员选择了一个嗜热微生物――海栖热袍菌的完全蛋白质组,该微生物有1877个ORF,其中46%没有赋予功能注释(Nelson等,1999)。在1877个目标中,有1376个(73%)表达克隆可以通过发酵和纯化过程导出来。这些克隆中的542个(约40%)能够产生微克级的可溶表达的蛋白质,并用于结构测定试验(Lesley等,2 ...
疾病相关基因的搜寻与壳隆的研究策略 当前疾病相关基因的搜寻与克隆的研究策略依然主要采用关联研究和定位克隆,其他如全基因组扫描、全基因组关联研究、候选基因关联研究等策略和方法都是从上述两种策略衍生出来的。另外,尚有基于染色体异常和动物模型等一些方法和策略。但值得注意的是所采用的策略方法并非孤立,而是相互借鉴,综合应用。随着对基因组特性和各种策略方法认识的加深、新的研究方法的出现以及分型技术的 ...
疾病相关基因的搜寻与克隆的研究历程 1.从功能克隆到遗传分析 20世纪40―60年代,许多研究生物化学和生物大分子的手段如电子显微镜、生物分子分离、提纯、蛋白质电泳相继出现,使人们在分子水平上研究疾病病因成为可能。许多酶缺陷的疾病就是利用这些方法将其基因确定的,如苯丙酮尿症等。功能克隆是基于疾病与正常之间明显可见的或直接与生化功能相关的线索确定与疾病相关的基因,主要是利用基因的产物蛋 ...
DNA变异 基因组DNA是人类遗传信息的载体,人类所有的生命活动均是通过基因经由其编码产物蛋白质来执行的。任何引起基因功能变化的突变(变异或体细胞突变)或表观遗传学(epigenetics)的改变必然对机体产生影响,甚至形成疾病。疾病相关基因的搜寻与克隆,其实就是寻找这些引起基因功能变化的突变或表观遗传学改变。找到了这些导致基因功能改变的突变或表观遗传学改变,也就找到了致病基因。这其中尤以 ...
SNP单倍型的检测 (1)分子单倍型分型(molecularhaplotyping):利用实验的方法直接获得单倍型。已有的技术包括等温滚环扩增反应(isothermal rolling―circle amplification)、碳纳米管检测(carbon―nanotube probing)、双倍体―单倍体转换方法(diploid to haploid conversionmethod)、 ...
大片段DNA拷贝数多态性 由于SNP在基因组丰富及与疾病和药物疗效等的相关性,备受关注。但在基因组中除了SNP和短片段插入(缺失)外,尚存在大片段DNA拷贝数多态性(长度约几十kb或以上)。受到净化选择(purlfFngselection)的作用,这些大片段DNA中的基因较少。至今已发现的受大片段DNA多态性影响的基因约有300个,其中有些基因与一些复杂性状(complextraits)或 ...
体细胞突变 除了可遗传的变异影响疾病的发病外,有时组织细胞在各种因素的作用下,其DNA也会发生突变,即体细胞突变(somaticmutation),使得该突变细胞的基因组不同于人体其他正常细胞的基因组。体细胞突变最常见于肿瘤细胞。在肿瘤细胞中,除了单个碱基的突变外,通常还涉及DNA大片段的缺失或扩增。如原发性肝细胞癌,户53等抑癌基因常因突变或缺失失活,而p―连接素(p―catenin)等 ...
定位克隆的核心是连锁分析技术 连锁是一种遗传现象,是指当同一条染色体上的某些位点由于相距很近,在减数分裂过程中这些位点发生交换的概率较小,所以就有较多的机会连锁在一起,从亲代传递到子代。位点间的距离越近,在减数分裂过程中发生交换的可能性越小,位点间重组的概率就越小;相反,两位点越远,则越容易产生重组体。这是利用连锁分析将致病相关基因从染色体中搜寻出来的定位克隆基础,为疾病基因定位提供了有力 ...
连锁分析法 连锁分析法主要适用于已知遗传方式单基因遗传病的基因定位,对具有复杂性状的多基因疾病进行遗传连锁分析时往往会受到多种因素的影响,如异位显性(epistasis)、不同发病年龄、不完全外显率、多个致病基因位点、遗传与环境相互作用、遗传模式、表现型比率及诊断不明确等,因此运用连锁分析研究多基因疾病受到一定程度的限制。针对连锁分析的上述限制性,目前采取的策略是基于血缘同一性的患者同胞对 ...
关于丁香通
公司信息
个人用户
企业机构


