

抗原设计
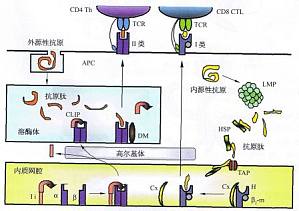
溶酶体途径--外源性抗原的降解 外源性抗原主要来自通过各种途径进入机体的非己成分,例如细菌产生的毒素。病原体所展示的PAMP由固有内体―溶酶体途径(endosome/ly~somepathway)加工提呈外源性蛋白质抗原大致分为四个阶段。蛋白质抗原加工提呈的两条主要途径1.MP:低分子质量多肽;Cx:钙联蛋白;TAP:抗原加工相关转运蛋白;Ii:Ia相关不变链Ⅱ类结合的不变链肽段。 ...

内源性抗原肽的加工 内源性抗原指胞质内出现的抗原,如内源性病毒、肿瘤抗原和某些自身抗原。与外源性抗原不同,内源性抗原主要通过胞质溶胶途径(cytosolpathway)完成加工提呈的过程。也有相应的四个阶段。 内源性抗原肽的加工 胞质溶胶途径对抗原的加工主要通过蛋白酶体(proteosome),一种存在于胞质溶胶中的大分子质量蛋白质水解酶复合物。动物细胞中蛋白酶体由20S的蛋白酶 ...
蛋白质降解的泛素―蛋白酶体途径 泛素(ubiquitin,Ub)是76个氨基残基组成的小分子多肽,可以以共价结合的方式与蛋白质的赖氨酸相连。蛋白质一旦接有泛素,称为发生泛素化(uhiquitylation)。泛素化在ATP的参与下被三种酶依序催化,形成蛋白质与一条泛素聚合链相结合的复合结构,进入蛋白酶体,然后降解为肽段(图8―15A)。此为生物大分子在胞质中降解的泛素―蛋白酶体途径(ubi ...
内源性抗原肽的转运 经蛋白酶体降解产生的内源性抗原肽必须进入内质网才能与I类分子结合,这一转运过程依赖抗原加工相关转运物(transporter associated with antigen processing,TAP)。这是一种位于ER膜上的跨膜蛋白,属ABC转运蛋白家族,由两个亚单位TAPl和TAP2组成,编码基因位于HLAⅡ类基因区中PSMB基因的近旁。每个亚单位反复穿越ER膜6 ...
MHCI类分子荷肽 I类分子α链和β2m在糙面内质网中合成后被转运到光面内质网。α链在到达内质网后立即与伴随蛋白结合。参与I类分子加工的伴随蛋白很多,主要的有Cx、热休克蛋白(HSP)和钙网蛋白(calreticulin)。 不同的HSP分子是以亚基分子质量(kDa)来命名的,例如,HSP70是亚基分子质量为70 kDa的蛋白质。目前发现的HSP已有近30种,分为4组,即HSP70 ...
内源性抗原的提呈 结合了肽的I类分子从内质网进入高尔基体,经糖化修饰后,通过胞吐空泡被转运到细胞表面,供CD8CTI-细胞识别。 内源性蛋白质抗原提呈途径的阐明对认识肿瘤抗原的本质有重要意义。肿瘤抗原多数由正常细胞基因突变而来,虽属非己成分,但其产生具有内源性抗原的特点。图8―17表明,突变基因产物经由胞质溶胶途径被I类分子提呈至肿瘤细胞表面后被CD8CTL所识别。这意味着肿瘤抗原 ...
MHC分子、抗原肽和TCR间的结合与相互作用 T细胞识别抗原的过程中,受体TCR和配体pMHC之间密切结合,而构成配体分子pMHC的两种成分即抗原肽和MHC分子之间,也发生相互作用。专门提出这一点之所以必要,是因为和通常出现的受体―配体相互配接(1igation)不同,此处涉及的是三种而不是两种分子,而且,更重要的是,三种分子都显示高度的变异性。因而,由此发生的相互作用既有普遍意义的一面, ...
蛋白质抗原加工提呈的两条主要途径 T细胞能识别APC表面由MHC分子提交的抗原肽,依赖于APC对蛋白质抗原进行处理和加工,并将抗原肽展示于细胞表面供TCR识别,称为抗原的加工提呈。 抗原加工与提呈分为针对外源性抗原和针对内源性抗原两条主要的途径。需要说明的是,内源性抗原并非自身抗原的同义词,外源性抗原也不等于非己抗原。内源性和外源性抗原的区分是根据它们在进入加工途径前所处的位置,即 ...
这些非典型可与上述两条主要途径或经典途径并存,使一种抗原可通过不同的途径被加工提呈,扩大了免疫应答的范围。但非典型途径往往出现在一些病理性条件下,在免疫耐受、抗胞内感染和抗肿瘤免疫中发挥作用。参与非经典途径的APC主要是DC和Mφ。蛋白质抗原加工提呈的非经典途径当内体/溶酶体因病理因素或药物处理而破损,外源性抗原肽可进入胞质溶胶,由I类分子提呈。 当L链结构异常 ...
抗原肽-MHC相互作用中等位基因特异性及其意义 上述结果已十分明确地显示,抗原肽和MHC分子间的相互作用能否实现,是由MHC等位基因分子抗原结合槽的结构特点、进入该结合槽的抗原肽是否符合其接纳抗原肽的共用基序,以及这些肽可能显示的抑制性残基的部位和类型所决定的。MHC等位基因的差异(多态性)体现在个体之间,因而,抗原肽-MHC相互作用的不同格局,有可能直接参与构成不同个体对同一抗原应答格局 ...

抗原肽的锚着残基和MHC分子接纳抗原肽的共用基序 抗原肽一般含有2个或2个以上与某个特定MHC分子结合的部位,称为锚着位(anchorposition),位于该部位上的氨基酸则称为锚着残基。这些锚着残基插入MHC分子抗原结合槽的小袋中,通过氢键与MHC分子相结合。抗原肽中间部位一般均有一定程度的隆起,可作为T细胞表位供TCR识别。 与同一型别MHC分子相结合的不同抗原肽,其锚着位和锚 ...
MHC I类和Ⅱ类分子抗原结合槽的结构特点 比较I类和Ⅱ类分子以各自的抗原结合槽接纳抗原肽的特点。这一特点仅仅体现了两类分子相互作用中的共性。MHC I类分子和Ⅱ类分子抗原肽结合槽的结构和相应的功能特点 I类分子α1和α2结构域各含4段β―片层和一个α―螺旋,共同组成I类分子的抗原肽结合槽,是氨基酸(α.α.)组成变异性最大的部分。该槽两端封闭,一般只能容纳8―10个氨基酸 ...
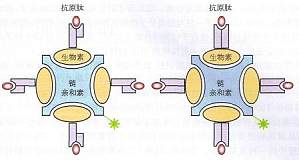
MHC四聚体 由于可溶性MHC单体分子与TCR的亲和力很低,解离快,而多价分子可与一个特异性T细胞上的多个TCR结合,使其解离速度大大减慢。为此Altman等提出借助生物素―亲和素级联反应放大原理构建MHC I类分子四聚体。该方法通过基因工程技术把长度为15个氨基酸残基的生物素酶底物肽(Bio A substrate peptide,BSP)加在MHCI类分子如HLA-A2重链的羧基端形成 ...
T细胞识别抗原中的MHC限制性 1974年Zinkernagel和Doherty首先证明受牛痘病毒感染的CBA品系小鼠(H-2k)中的CTL只能杀伤H-2单元型相同的病毒感染的靶细胞,而不能杀死同一病毒感染的MHC背景为H-2b的小鼠细胞。这是首次揭示T细胞识别细胞表面抗原肽的同时还需特异性地识别提交抗原肽的MHC等位基因分子。以后证实,不仅CTL-靶细胞间,而且M旷Th及Th-B间的相互 ...
应用共用基序寻找和获取带有T表位的特异性抗原 对特定抗原或病原体的易感性如果受MHC等位基因的约束而呈现出一种称为疾病与MHC关联格局,可以通过该MHC等位基因分子抗原结合槽的共用基序,寻找特异的致病性抗原表位。如果某一种I类分子接纳9肽时的共用基序,在锚着位p2和p9的氨基酸残基为脯氨酸(P)和亮氨酸(L)(注意:人和小鼠主要MHC等位基因分子的共用基序已经公布,可以查找),则可以用电脑 ...
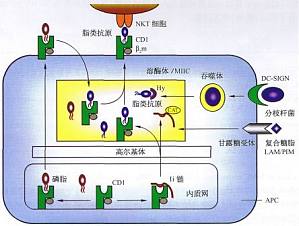
CDl分子对脂类抗原的提呈 CDl分子在结构上与MHC I类分子相关,但Cm基因位于MHC外,无多态性。人类有五个紧密连锁的CD5基因,四个表达。编码蛋白分成两组,第一组包括CDh、CDlb和CDlc,CDld属于第二组。CDlb和CDlc提呈的脂类抗原,主要来自分枝杆菌胞壁成分,包括糖脂和磷脂等,如霉菌酸、葡萄糖单霉菌酸脂、脂阿糖甘露聚糖等。CDld分子能提呈疏水肽,也能提呈某些脂类抗原 ...
免疫受体酪氨酸激活基序 除了蛋白质磷酸化,T细胞信号转导的实施,还需要把各种游离于胞质中的激酶和信号分子招募到胞膜内侧和受体分子近旁,为信号转导创造条件。其中发挥关键作用的结构,主要是受体(或受体相关分子)胞内段上特定的ITAM、识别这些基序的SH2结构域,以及带有SH2结构域的激酶和衔接蛋白。 已知T细胞受体相关分子上ITAM的结构为…D/Exx I x(7-11)I,…其中以方 ...
免疫突触引起跨膜分子及信号转导成分的多聚化 T细胞表面免疫突触的形成,不仅增加TCR与pMHC间的亲和力,并引发胞膜相关分子的一个重要物理变化--多聚作用(multimerization),使参与T细胞激活的各种跨膜分子如TCR/CD3、辅助受体CD4(或CD8)、CD45等相互靠拢成簇(clustering)。 同时,多聚作用引起受体关联性酪氨酸激酶,主要是非受体型SrcFTK家 ...
CD45分子参与启动信号转导 然而事实上,上述过程并不产生。Src分子有两个酪氨酸残基位点,一个位于催化结构域即SHl内394位置处,是激酶显示活性的关键部位,也是相互磷酸化中Src分子间磷酸根转移的靶目标。另一个酪氨酸残基在Sre分子C端505位置,发挥抑制作用。在未激活的T细胞中组成性表达一种称为Csk的蛋白酪氨酸激酶(图9―1)。Csk指C端Src激酶(C―terminal Srck ...
MAP激酶相关途径 1.Ras蛋白与信号转导 原癌基因FaS产物p21ras的参与。p21ras(简称Ras蛋白)分子质量为2lkDa,属鸟苷酸结合蛋白,是小G蛋白家族中的一个重要成员。G蛋白分两类,除了此处的小G蛋白外,另一类为异源三聚体G蛋白(heterotrimerlc G protein)。异源三聚体G蛋白相关受体为七次跨膜型结构,是一个包括100多个成员的大家族(见第五章图 ...
关于丁香通
公司信息
个人用户
企业机构


