

抗原设计
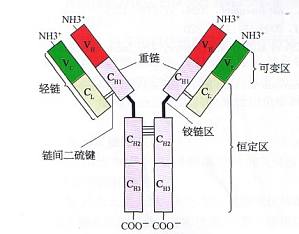
免疫球蛋白(immunoglobulin,Ig) 免疫球蛋白(immunoglobulin,Ig)是血液和组织液中的一类糖蛋白,由B细胞接受抗原刺激后增殖分化为浆细胞所产生,是介导体液免疫的重要免疫分子。近年研究证实,Ig与抗体(Ab)在结构及功能上完全一致,因此可认为二者的概念等同。Ig可分为分泌型(secretedlg,SIg)和膜型(membranelg,mlg),前者主要存在于血液 ...

免疫球蛋白的J链和分泌片 所有Ig分子都是由两条重链和两条轻链组成,但某些类型的Ig还含有其他辅助成分,分别是J链(joiningcham)和分泌片(secretorypmce,SP)。免疫球蛋白的J链和Ⅲ结构 J链是一富含牛胱氨酸的多肽链,由浆细胞合成,主要功能是连接单体Ig分子使其成为多聚体。 IgA的二聚体和IgM的五聚体均含J链IgD和IgE常为单体,不含J链。 SP为一 ...
免疫球蛋白水解片段 Ig分子中有些部位在一定条件下,容易被蛋白酶水解为各种片段。常用的蛋白酶有木瓜蛋白酶(papain)和胃蛋白酶(pepsin)。 Ig酶解的片段获得,大大促进了Ig分子结构和功能的研究,不但加深了对Ig分子的了解,同时也为各种基因工程抗体的构建打下了基础。如DNA操作和抗体改型技术的发展,使得人们可根据抗体水解片段的结构和功能关系,产生更多有功能的Ig改型分子片 ...
免疫球蛋白的识别和结合抗原功能 1.Ig特异性识别和结合抗原的活性由其V区CDR所组成的抗原结合槽结构所决定的。抗原结合槽和抗原表位的结合是空间构象上的互补,结合小型抗原如半抗原或短肽的抗原结合部位大多为袋型或沟槽型,对大型抗原如蛋白质,大多为平面型。抗原结合槽结合抗原可以是锁―匙(1ock-key)样的直接互补,也可是结合抗原表位后构象诱变而致的诱导吻合(inducedfit)。由于Ig ...
各类免疫球蛋白的生物学特性 1.IgG 血清和胞外液中主要的抗体成分,约占血清Ig总量的80%(表3-2)。kG白出生后3个月开始合成,3―5岁接近成人水平,半寿期为20~23d,是再次体液免疫应答产生的主要抗体,亲和力高,在体内分布广泛,具有重要的免疫效应。IgGl、IgG3、IgG4可穿过胎盘屏障,在新生儿抗感染免疫中起重要作用;IgGl、IgG2、IgG4可以其Fc段与葡萄球菌蛋白 ...
免疫球蛋白编码基因及蛋白合成 Ig轻链和重链及其C区和V区分别由分布于不同染色体的多个基因片段所编码(表3―3)。重链的胚系基因中有许多基因片段,包括众多的V基因片段、多个D基因片段、几个J基因片段和5类C基因片段;轻链的胚系基因中也有许多基因片段,包括V基因片段、J基因片段和C基因片段,但不含D基因片段。肚链Ig基因的染色体定位 由于编码轻重链可变区的基因都有V、J片段,为了分 ...
免疫球蛋白胚系基因结构 1.人重链胚系基因结构 人重链基因定位于第14号染色体长臂,跨度约1 100kb,由V、D、J和C四种基因片段组成,包含约95个Va基因片段(分VHl一VH7七个家族,其中功能性基因片段65个)、27个D基因片段、6个JH基因片段和9个CH基因片段(表3-4)。Va基因片段位于上游。D基因片段位于VH和JH基因簇之间,JH基因位于DH下游,与下游C基因区相隔7Kb ...
免疫球蛋白胚系基因重排的机制 RSS包括七聚体(heptamer)和九聚体(nonamer)两种核苷酸序列,前者为CACAGTG,后者为ACAAAAACC。七聚体和九聚体之间含一间隔序列(spacer),分别为12bp或23bp长短。这种“七聚体―间隔序列―九聚体”结构即为重组信号序列。在重组酶识别和作用下,带有12bp间隔序列RSS的基因片段只能和带有23bp间隔序列的片段相结合而发生重 ...
免疫球蛋白编码基因的转录和翻译 虽然所有的体细胞均含有Ig胚系基因,但只有B细胞的Ig胚系基因经重排后才能转录、翻译和表达Ig多肽链。B细胞受抗原刺激后,经过一系列信号分子的作用,激活细胞内相关的转录因子,在转录调控元件(如启动子、增强子)和调控因子(如调节蛋白及转录因子)的作用下,H链和l链基因独自进行转录。但在一对同源染色体上的重链或轻链基因只有其中的一条染色体上的/g基因得到转录和表 ...
免疫球蛋白编码基因的二次重排 其中会发生一种称为受体编辑的现象。受体编辑(receptorediting)又称二次重排。在Ig胚系基因重排过程中,VDJ连接可发生读码框架位移,出现终止密码子而导致无功能产物的出现。过去曾认为一个分化中的B细胞在两条染色体的重排都失败后或重排产生自身反应性Ig受体克隆时B细胞发生凋亡。但后来发现一次重排失败后可发生二次重排,形成替换基因(replacemen ...
免疫球蛋白的多样性及其产生机制 Ig胚系基因通过基因重排和抗原的刺激,由活化B细胞分化发育为浆细胞后产生Ig。自然界中有数量巨大的抗亨物质,可刺激机体产生同样数量巨大的特异性免疫球蛋白。由此构成了免疫球蛋白多样性(1gdiversity)。 Ig有两个基本的特哇:作为抗体可特异性结合抗原,但又可同时作为抗原分子,诱导机体产生抗免疫球蛋白的免疫应答。虽然Ig分子的基本组成为H2L2, ...
体细胞高频突变形成免疫球蛋白的多样性 体细胞高频突变发生于基因重排后成熟B细胞受抗原刺激后的分化发育阶段,而并非像上述两种机制发生于胚系基因片段上,突变频率高,而且只出现于次级淋巴器官的生发中心。主要方式是替代性点突变。点突变的位置并非完全随机分布,主要发生于重排后的可变区基因3,和5,端300个左右的碱基内,其突变频率较细胞中正常基因突变频率高105倍,这样既能产生最大序列的多样性,又不 ...
基因工程抗体 DNA操作技术和基因工程技术的发展,使得人们不仅可用杂交瘤技术生产单抗,也可通过基因工程手段人为制备各类抗体。抗体可在动物细胞中表达,如骨髓瘤细胞(myelomacell),结合大规模培养的生物反应器,抗体产量据报道可高达500mg/Lo也可用其他的动物细胞来表达,包括昆虫细胞在内。 细菌系统因其遗传背景清楚、转化效率高、繁殖迅速、培养基便宜、而且早已建立的微生物发酵 ...
抗体片段及其衍生形式 1.双特异性抗体 将两套轻链、重链基因导入骨髓瘤细胞中,选择合适的抗体恒定区及Ig类型,可获得产量大、均一性和高纯度的双特异性抗体。另外,以化学交联技术或杂交―杂交瘤技术也可获得双特异性抗体。 2.Fab抗体 Fab抗体是由重链Fd(即VM+CHl)和完整轻链通过二硫键结合而成的异二聚体,仅含一个抗原结合部位。将重链Fd和完整轻链的编码基因连接,5,端融合 ...
主要组织相容性复合体 主要组织相容性复合体(MHC)是由一组高度多态性基因组成的染色体区域。MHC基因产物能在不同细胞表面表达,通常称之为MHC分子或MHC抗原,又因这些抗原在器官移植中代表供受者双方的组织相容程度,也称为移植抗原或组织相容性抗原。脊椎动物中,从鱼到人类都存在结构与功能相似的MHC遗传区域,如小鼠的MHC是H-2,堵的MHC是SLA,人的MHC是HLA。 MHC是现 ...
小鼠H-2(小鼠的MHC复合体)系统 1900年Landsteiner发现人体ABO血型,引起了寻找各种动物红细胞抗原的热潮。1939年Gorer在鉴定近交系小鼠血型抗原时发现四组血细胞抗原,后证实第Ⅱ组抗原与肿瘤及移植物的排斥密切相关,就将之重新命名为H-2即histocompatibility-2。因而现今,H-2代表了小鼠的MHC复合体。 H-2复合体位于小鼠第17号染色体的一 ...
扩展的主要组织相容性复合体 xMHC这段区域长度从HLA工类区域最远端的HCP5P15算起,最终扩展到HLAⅡ类区域的接近着丝粒处的KIFCI,共7.6Mb,包括下列5个亚区及421个基因座位。 (1)扩展的I类亚区(extendedclassⅠsubregion):长3.9 M山含有182个基因座位。 (2)经典的I类亚区(classicalclassⅠsubregi。n) ...
人类主要组织相容性复合体HLA HLA(human leucocyte antigen)基因系统是人类主要组织相容性复合体。人类第一个HLA抗原由法国Dausset在1958年从三个多次接受输血患者的血清中检出,当时命名为MAC,相当于目前正式命名的HlA―A2加上HLA―A28。基因产物称为HLA抗原或白细胞抗原。有关HlA的研究工作在临床器官移植配型和国际大协作推动下发展十分迅速,涉及 ...
HLAⅢ类基因区 位于HLAI类和Ⅱ类区之间从PPIP9到BTNL2包含有62个基因座位.长度为0.9Mb。其中58个表达基因,2个假基因,每14 516bp就有一个基因,因此是整个基因组中基因密度最大的一个区域。除了LOCA01250基因外,其余基因均在1999年已发现。此外,在Ⅲ类基因区的表达基因中,有的产物发挥重要作用.如参与细胞转录调节(CREBLl、BTKl9、PBX2、RDBP ...
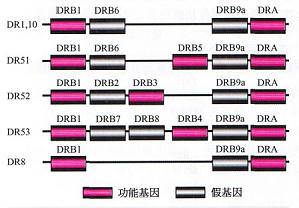
HLAⅡ类基因区 HLAⅡ类基因区位于HLA复合体的着丝粒端,即从DRA到HLA-DPA3的一段区域,长度为0.9Mb,包含34个基因座位,其中16个为表达基因,3个后选基因,15个假基因。在16个表达基因中除了BRD2(原名RING3)外均有免疫学功能。Ⅱ类区域内至少含有DR、DQ、DP、DOA、IX)B和DM六个亚区。 (1)HLA-DR亚区:DR亚区有1个DRA基因及9个DRB ...
关于丁香通
公司信息
个人用户
企业机构


