

数据库

人类的水分由机体蒸发,除发汗外,还可以由皮肤和呼吸道粘膜进行,后两者称为不感蒸发。不感蒸发是一种不间断的基本水分的损失,与体温的恒常性的维持机制(如发汗)是有区别的。皮肤的不感蒸发是表皮细胞间隙中组织液的水分直接透过皮肤而蒸发掉,所以在缺少汗腺的动物也可发生。人在通常的环境中,安静时的不感蒸发量,为每1平方米体表面积每小时约30克。 ...
即不会遗传的变异,是由于环境变化所引起的变异和所谓获得住状都属于不遗传的变异。 ...
在生物对某一刺激发生反应后,在一定时间内,即使再给予刺激,也不发生反应。一般称此期间为不应期。例如给神经肌肉等应激性器官或部位以刺激,刚产生兴奋之后,再给予第二次刺激则无效。该时期被认为是该器官或部位是处于兴奋状态的恢复时期。不应期分为:不论第二次刺激强度多大均无反应的绝对不应期(absolute refractory period);以及以后给予强刺激则可能发生反应的相对不应期(relati ...
发生感染的溶原噬菌体基因组不插入寄主细菌的染色体中,而且受自身形成的阻遏物的抑制不能复制增殖,称这种状态为不育溶原化。这类噬菌体基因组在细胞分裂时只进入子代细胞一方,因而就形成只有一个子代细胞中有噬菌体的基因组,其它大部分都成为不具有的无性繁殖系。这种遗传方式称为单线遗传。 ...
借助细菌噬菌体转导到细菌中的染色体片段,既不插入寄主染色体中,也不增殖,但仍表现出机能,这种状态称为不育转导。它是一种特殊形式的转导,被转导的基因在每次细胞分裂时只进入子代细胞。这种情况可以在营养缺陷型转导或鞭毛形成性状的转导中观察到。 ...
1 指不产生发育成下代植物种子的这一特性。广义的不育性包括因环境条件而不开花或过早落花的,以及种子不具发芽能力的,或是像白化苗那样能发芽但无繁殖能力的。狭义的不育性,则是指从生殖细胞形成直到受精,或是作为担负这一进程的生殖器官的机能、形态、位置,以及从合子到发育成种子这一整个过程,基于某种原因,而不能产生后代的现象。M.B.Crane(1929)把因生殖器官退化而产生的不育性,称为形态性不育性 ...
一般遗传重组是在染色体相同部位之间对称地发生交换,而不正常重组是指在染色体不相同部位之间所发生的交换。转导噬菌体的形成就是由于不正常重组的结果,它们发生重组的位置是不一定的。不正常重组发生的频率虽很低,但它却是染色体上各部位基因发生缺失或重复的可能原因。 ...
在日本本州岛北海道岛间所划的生物界线,又称津轻海峡线。系1880年布莱基斯顿和普赖尔(T.W.Blakiston,H.Pryer)根据鸟类分布所提出的界线,由米尔恩(J.Milne,1883)命名,最初确认是以属于中国东北亚区的本州与属于西伯利亚亚区的北海道之间为分界线,但后来根据在秋田八幡平也发现了北海道产的熊,以及根据其他鸟类和淡水鱼等情况作了修正,因此现在对这条线的评价就不象从前那样高 ...
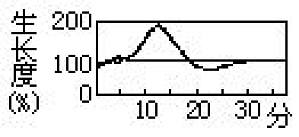
把闪光须霉(Phycomyces nitens)的孢子囊柄和一种向日葵的胚轴先放在暗处,用不同强度的光作很短时间(如四秒钟)的照射,然后在较短时间后检查光对植物体生长的影响。结果发现须霉有如图所示的情况:先是促进其生长(一次反应),然后生长比在暗处时还慢以至达到最低生长(二次反应),而向日葵则与上述情况完全相反,起初生长缓慢,然后则表现生长有被促进的倾向。植物的这种在暗处生长速度随光照急剧改变 ...

鬼伞等属担子菌的菌丝间的特殊接合过程,当单倍的初生菌丝与异核体的次生菌丝发生接合时,初生菌丝接受次生菌丝中的核而变成次生菌丝的现象称为布勒现象。如果初生菌丝进行体细胞接合,一般由异核的,即有双核的细胞构成次生菌丝,而这种次生菌丝当和初生菌丝接合时,又再度发生体细胞接合。假如构成次生菌丝对的两个核之一和初生菌丝的核有亲合性,则该核即进入初生菌丝中而使其次生菌丝化。 根据初生菌丝和次生菌丝间的性的差 ...
H.M.Bruce(1959)发现,交尾后的雌性小白鼠立即与其他雄性同居时,则不妊娠,此现象称为布鲁士氏效应。据认为这是因为雄性尿中的信息素能抑制其黄体的活性化,但是因为有个体识别的关系,所以还不能单纯用信息素的作用来说明这复杂的反应。 ...
引起羊、牛、猪等家畜传染性流产症的一属病原菌,为D.Bruce(1887)发现。是直径约0.6微米的球菌或长度在1微米以上的杆菌。革兰氏染色阴性,不运动,好氧性,仅可利用少量的碳水化合物,而不形成酸。可在肝浸出物等动物性培养基中生长,产生大量氨和硫化氢。最显著的特征是培养中需要二氧化碳,在10%的CO2中能进行良好的发育。所产生的毒素和痢疾菌等肠道细菌相类似,被提纯的一种菌体抗原是糖脂-糖肽复 ...
存在于十二指肠壁深部的腺体,亦称十二指肠腺(duodenal gland)。在胃的幽门端,有与此构造和机能相类似的腺体。分泌液呈碱性,腺细胞由粘液细胞所构成。 ...
光化学的效应是吸收的光强度与照射时间的乘积成比例的法则,由德国的R.W.Bunsen与英国的H.E.Roscoe两位化学家(1855—57)创立的。J.Loeb(1918)举出很多例子〔藤壶的无节幼体(naupli-us)等〕,证实这个法则适用于动物的趋光性反应。并作为他的趋性学说的一个根据:趋性完全是机械控制反应,以后还确认这一法则也适用于植物体的向光性与动物的各种感光反应(例如海鞘水管的收 ...
棘皮动物,如海胆,沿许多管足分布的方向有五个辐状带,其中间又有棘将五个带分开。前者叫步带,有整齐的两列步带板(ambulacral plate),其上有连通管足的小孔规则地排列着。在步带和步带之间是间步带,在间步带板(interambulacral plate)上,有大大小小的突起即棘附着点。有步带的部位,是体的主对称面,沿此方向有水管系的辐步管、血窦系的辐血窦、辐神经等在体壁下走行.间步带位 ...
海盘车类腕下面中间从口至腕端走行的一条沟。左右各有一对步带板并排成屋脊形。贯穿步带板,生有成列的管足,所以又称为管足沟。
也称围口水管(circumoral water vessel)或环水管(ring water vissel,circular water ves-sel)。是在棘皮动物水管系统中其围绕在咽部的环状部分。由此引出5条辐步管达到反口极,此外还附有波里氏囊和铁特孟氏体(海星类)经过一条石管由体表的多孔体开口于外界。 ...
简称部分变性图或变性图.一般来说DNA的双链结构的稳定性取决于它的碱基组成,A-T含量高的部分其氢键在比较低的温度中就会拆开,所以容易变性。如把分子内部的碱基组成有不相称的DNA放在中等的变性条件下之后,用甲酰胺等来固定单链部分,在电镜下进行观察,根据所显示的变性部分的位置和长度就可以作成部分变性图。变性部分的分布和长度等特征可以作为以后电镜观察该种DNA时的标记之用。此外还可以从部分变性图上 ...
只有一部分基因组发生加倍的二倍体。特别是细菌的杂合体,这种状态很普通,由受体菌的完整染色体与供体菌的染色体片段形成部分二倍体。另外,利用转导噬菌体或F′因子将供体菌的染色体片段导入受体菌中;也可以维持稳定的状态。 ...
通过半融合形成的部分合子。特别是细菌的杂交中,Hfr菌株的染色体通常并不全部转移到F-菌株中,而只移入一部分,因此,合子中F-染色体(此处称内基因子)和部分Hfr染色体(此处称外基因子)经过半合子时期后,相同部分发生基因重组,最终形成重组体。另外在F′和F-之间杂交时,F′染色体(附着了一部分细菌染色体的F因子)进入F-菌,以自主的质体形式存在着,因此可以得到部分二倍体菌。在转导时,有时转导噬菌体 ...
关于丁香通
公司信息
个人用户
企业机构


