


最初是通过琥珀干馏获得的,所以有琥珀酸这个名称。除广泛存在于植物界外,还可由细菌发酵产生。在动物组织中也有可形成和转化成琥珀酸的强有力的酶系存在,它是三羧酸循环中的一环,在琥珀酸脱氢酸的作用下,通过细胞色素系的电子传递而形成延胡索酸。
系催化琥珀酸转化为延胡索酸的脱氢反应的酶( E. C. 1. 3. 99. 1)。生理的电子受体还不太清楚。属于黄素酶类,每约20—30万分子量中含有1个FAD,CH2—COOH HOOCCHFAD和酶蛋白以共价键结合。酶中尚含有数个原子非铁血红素。Fg(CN)63-和吩嗪(phenagine)是它有效的人工电子受体。是巯(SH)酶的一种。可受丙二酸和草酰乙酸的拮抗性抑制,此酶存在于线粒体内膜 ...
用氧来氧化琥珀酸为延胡索酸的酶类。琥珀酸脱氢酶稳定地存在于线粒体内膜上,不能以可溶性的形态被提取,与细胞色素等密切结合,形成将电子传递给氧的复合酶系,因此很早以来就对琥珀酸氧化酶系进行了研究。作为成分,尚有FAD、非铁血红素、辅酶Q、细胞色素b、c、c、a、铜。氧化1分子琥珀酸,可从 ADP和无机磷酸偶联生成二分子的ATP。 ...
简写为am。无意义突变的一种。因硷基置换而产生的突变,使对应于某种氨基酸的密码子变为琥珀密码子UAG。这是以最初与埃皮施泰因和施泰因贝格(R.H.Epistein和C.M.Steinberg)一起从T4 噬菌体中分离这种突变株的贝尔施泰因(H.Bernte- in)的名字来命名的。因为德语Bernstein(琥珀)的英译为amber,所以称这种突变为琥珀型突变。 ...
CoA-S-CO-CH2CH2COOH。 CoA的SH基与琥珀酸形成硫酯键,为α-酮戍二酸通过α-酮戊二酸脱氢酶氧化时生成。是三羧酸循环的组成环节之一。α-酮戊二酸 CoA NAD 琥珀酰CoA NADH H CO2 心肌的粗制酶,因含有琥珀酸CoA脱酰基酶,而水解生成琥珀酸,但琥珀酰CoA合成酶起作用时,可将高能键转移而生成GTP琥珀酰CoA GDP 磷酸琥珀酸 GTP CoA 这是基质 ...
侧重于某一个性状,此性状只有在二个以上的非等位基因共存时才会表现出来,这种由于互相补充而使一个性状得以表现的非等位基因,称为互补基因。典型例子是W.Bateson、E.R.Saunders以及R.C.Punnett(1906)研究过的决定香豌豆花色的基因,使不同的二种白花品系杂交,结果F1花呈紫色,而F2中紫花与白花以9∶7的比例分离。这是因为二个互补基因(形成色素原体的基因和形成显色酶的基因)分 ...
这是研究二个突变是否属于基因的同一功能单位的方法。通过这种测定,能检测出顺反子和互补子。互补测定最早曾由E.B.Lewis(1951)用于果蝇,以后S.Benzer(1957)把这一方法应用于噬菌体,并提出了顺反子的概念。所以这种测定法又称为顺式-反式测定。当二个突变位于同一细胞内的同一染色体或基因组上时,此排列称为顺式排列(cis-arrange- ment, cis-configurati ...

二条多聚核苷酸链依3′5′和5′3′的方向排列时,按照腺嘌呤对胸腺嘧啶(或尿嘧啶),乌嘌呤对胞嘧啶这样的碱基配对原则所成的顺序,称为互补碱基排列。双链DNA整个分子上都存在互补的碱基排列,而在单链的tRNA和RNA噬菌体的RNA等分子内,具有互补碱基排列的区域可部分地形成双链结构。 ...
通过互补测定绘制互补图时所检出的顺反子内的遗传功能单位。亦称互补单位(complementatio- n unit)。属于不同互补子的二种突变型处于反式排列时,可显示顺反子内的互补性。也能同时影响二个或二个以上的互补子的突变。构成一个顺反子的互补子数目量是一定的,一个顺反子普通由几个互补子所构成。但已知有的顺反子是由10个以上的互补子所构成。一般认为互补子相当于多肽链的折叠起来的一个片断的基因部位 ...
分离属于同一顺反子的许多等位基因突变型,在这些突变型之间进行互补测定,可得到顺反子内显示互补的组合和不显示互补的组合。把显示同样类型互补性的突变型作为一群,对显示互补的突变型用线段来表示,不显示互补的突变型则用重叠的线段来表示,而用图来表示这些突变型在这个顺反子内的互补关系。这种图称为互补图,在这个互补图中顺反子内能够区别的最小单位称为互补子。在互补图上互补子的排列一般呈直线,但也有呈圆形的。 ...
同一细胞内含有二种突变基因组时,可以看到各突变体原所没有的性状表现出来或有的功能得到促进,这时两个突变体表现为互补,这个现象就称为互补作用。互补作用可通过互补测定来进行研究。也就是两种突变以各种方法表现于反式排列时,也就是当顺式排列时若产生与野生型一样或者近似于野生型的表型,说明这二个突变表现了互补作用,这两个突变属于不同的顺反子。这种互补性有时亦称为顺反子间的互补作用(intercisero ...
为共生的一种形态,指处于共生的双方,互相都能从对方得到某种生活上的利益,此称为互惠共生。双方得到的利益性质不一定是相同的。也有一种见解认为,互惠共生内容甚广,甚至不经常接触的关系也包括在内,几乎与互惠作用作同义语用。但一般多数是指经常的共同生活是双方不可缺少的情况而言,与此相反,双方并不是必须的互惠共生,称为原始共生。共生(symbiosis)一词,其原意是指广泛的共生,但在互惠共生中特别是限 ...
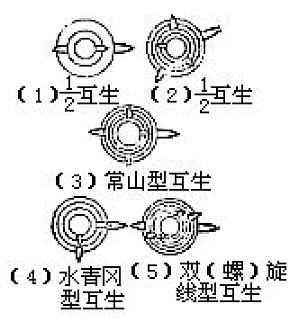
为叶序的一种形式,即茎的每节着生一片叶时,不论其如何排列,均称为互生叶。螺旋状排列的叶序是最普通的互生叶序,广泛见之于营养叶、花序和球果等。但也有周期性出现的几种不同开度的,如常山型(Oxixate type)叶序(180°、 90°、 180°、 270°的周期为4列纵生)、水青冈型(Fagus type)叶序(90°、270°的周期成特殊的二列纵生)。此外,还有总和不足360°,两种不同开 ...
俄国克鲁泡特金(ПАкропоткин)观察到在西伯利亚草原的高等脊椎动物,在严冬是通过互助来克服严酷的自然条件的,认为动物界中存在着互助的规律,从而否定了生物界和人类社会中生存竞争是普遍规律的说法(1902)。提示进行这种观察的是俄国动物学家凯斯列尔(ккесслер)。也有人认为,互助只不过是自然淘汰的一种形式而已。 ...
这是华莱士(A.R.Wallace,1868)提出的生物地理学上的著名界线。它从巽他群岛的巴厘岛和Lombok岛之间的狭长海峡,北上经过婆罗洲及Cele- bes之间的马卡塞(Nacassar)海峡,又自棉兰老岛南部延至东部。这条线以西连接马来半岛而位于一个大陆架上,各地具有共同的东洋亚区(旧热带区)的生物,而这条线以东,全部属于澳洲区。特别是巴厘岛与Lombok岛之间的狭长海峡,成为连接两大 ...
1953年华森(J.D.Watson)和克里克(F.H.C.Crick)关于DNA(脱氧核糖核酸)的分子结构提出的模型,主要根据威尔金斯(M.H.F.Wilkins)和富兰克林(R.E.Franklin)关于 DNAX射线衍射的资料,以及 E. Chargaff对来自各种生物的DNA样品进行碱基组成分析的结果,并基于腺嘌呤和胸腺嘧啶,鸟嘌呤和胞嘧啶(或其衍生物)之比分别为1而设计出的模型。按此 ...
指种子植物进行有性生殖的枝条,是由相当于茎的花轴和相当于叶的花叶构成的。花叶分化为花被(萼片、花瓣)、雄蕊和雌蕊几部分,也有把小苞包括在内的。虽然G.Gung和F. Wolff也这样把花看作是花轴和花叶构成的。但是自从哥德(G.W.von.Geothe)提出变态学说以后,这才为一般人所承认。作为花的最古老形态,在针叶树型的球状花,可上溯到石炭纪的果鳞木(Lepidocarpon),在苏铁型的大 ...
为构成花冠的不育花叶。花瓣与萼片不同,一般其基部细小,状似爪(claw),除此以外的部分称为瓣片(limb)或叶片(blade)。与普通叶相比,其组织分化一般较为简单,维管束一般细小,分支少。主脉不明显。另外在花瓣上未发现有叶绿素,代谢活性低。 ...
不育花叶没有花萼和花冠的区别时,其总体称为花被,而其组成叶称为花被片。花被多见于单子叶植物Tuliea gesneriana)、天香百合(Lilium auratum) 。根据花被片的序列和形态区别为内、外二轮时,外侧的称为外花被,内侧的称为内花被,一般大小、颜色和形态多少有些不同。百合在三枚外花被片之间有三枚内花被片。花被一词,广义上也可说是花萼和花冠的总称。在此情况下,花萼和花冠具有明显区 ...
为以花叶的种类和数目作为花组成的表示式。是由A.H.R.Grisebach(1854)引入植物学的,所以一般都遵从德国式,即以K表示花萼,C表示花冠,P表示花被,A表示雄蕊群,G表示雌蕊群,如东北堇菜(Viola mandschurica)的花程式为↓K5-C5A5G),郁金香(Tulipa gesneriana)的花程式为★P3 3A3 3G,桔梗(Platycodon grandi-flo ...
关于丁香通
公司信息
个人用户
企业机构


