


生物安全实验室气流流速流向和气密性检测 1.气流流速的检测 (1)单向流气流测定方法:距送风面0.5m的垂直截面。截面上测点间距不应大于2m,测点数应不少于10个,均匀布置。仪器用热球风速仪。 (2)评价标准;应符合生物安全实验室设计的每小时换气次数换算的气流流速,同时应考虑实验室内气流流速不应0.24m/hr。 2.气流方向应按以下要求进行检测和评价 (1)测定方 ...
免疫系统的个体发生与系统发生 免疫系统的个体发生和系统发生遵循从非特异性免疫到特异性免疫的规律。机体出生后即出现固有免疫,适应性免疫须待出生后接触抗原才能产生。免疫系统的进化中,最早出现的也是非特异性免疫,而淋巴细胞启动的适应性免疫,属免疫系统进化的高级形式。 吞噬细胞参与的固有免疫,在无脊椎动物的原虫和海绵中已经出现;NK细胞和淋巴细胞行使功能则在脊椎动物出现之后;MHC再要晚一 ...
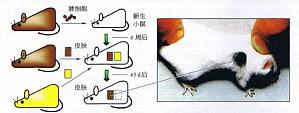
免疫学范畴的自身和非己的分辨 1.什么是“自身” 生物学中的定义是:机体胚系基因(gene in germ-line)的编码产物。免疫学中还要加一条,机体免疫系统发育过程中遭遇过的物质。 根据后一定义,属于“非己”的外来成分一旦接触未成熟的免疫系统,有可能被视为“自身”。例如,向尚未完成免疫系统发育的新生小鼠输入同种异型细胞,受者免疫系统会将该细胞表面的非己抗原视为自身。小鼠长大 ...
免疫系统的功能 作为生命科学中的一门独立学科,免疫系统履行下列三项功能: 1,免疫防御(immunedefense) 主要指抗感染,这仍是免疫系统应该担负的最重要的功能。不仅因为入侵机体的病原体种类繁多,包括细菌、病毒、真菌、支原体、寄生虫等,还因为会有新的病原体出现,并对人类和动物造成危害。 2免疫监视(immunesurveillance) 主要指识别和清除新出现的 ...
抗原表位 免疫细胞通常难以识别整个抗原分子,而仅识别抗原大分子上的一个特定的部分。称为表位(epitope)或抗原决定簇(antigenicdeterminant)。因而表位代表了抗原分子上的一个免疫活性区,负责与免疫细胞表面的抗原受体或游离的抗体分子相结合。严格说来,抗体的特异性是针对表位而不是针对完整的抗原分子。T、B细胞往往识别抗原分子上的不同表位。抗原分子以其B细胞表位与抗体分子 ...
决定免疫原性的抗原因素 机体免疫系统能否对抗原起反应,涉及构成抗原的各种大分子及其特性:而抗原进入机体是否诱发有效的特异性免疫应答,还取决于机体的多种生物学特性。因而免疫原性的强弱既由抗原因素决定,也与机体的生物学因素有关。 抗原因素 (1)异物性:这是对非己成分的另一种提法。自身当》己的区分,首先是后者具有异物性和外源性。而且,外源性的强弱和物种间胚系基因的差异程度有关。抗原 ...
固有免疫和适应性免疫对抗原的识别 外来抗原(如病原体)进入机体后,最早出现对抗原的识别和应答发生在0-4h:在该时相中,参与免疫系统识别非己成分的是一些现存的效应分子,如溶菌酶、C反应蛋白、甘露糖结合凝集素、血清淀粉样蛋白、血纤蛋白原等,甚至是一些可与病原体起反应的预存抗体。然后进入早期诱导性快速应答时相(4―96h),在抗原的动员和激发下,各种参与固有免疫的效应细胞如NK细胞和Mφ被激活 ...
决定免疫原性的生物学因素 (1)宿主的遗传背景:不同MHC背景的实验动物对同一抗原产生的应答格局可以有明显差异,由此发现了调控特异性免疫应答的免疫应答基因(Irgene),并确认Ir基因就是某些特定的MHC基因或等位基因。 (2)引入抗原的剂量和途径:抗原剂量必须适当,过高或过低将导致免疫无应答或免疫耐受。在数周内反复注射同一抗原比一次性注射效果好,因为可有效地激发抗原特异性淋巴细 ...
T细胞表位的特性 表达T细胞表位的抗原必须有可提呈性,因而表位的形成涉及APC和靶细胞对抗原的加工处理。此时,原有抗原分子是否保持其构型并不重要,也不会影响T细胞表位的下列特性。 (1)经抗原加工产生的抗原肽,参与构成T细胞识别中的“MHC抗原肽-TCR"三分子复合物,简称三元体。 (2)三元体中进入MHC分子抗原结合槽中的小肽,通常以其两端的锚着残基和MHC凹槽内壁结合, ...
B细胞表位的特性 (1)由抗原表面的亲水性氨基酸残基组成,易于接近BCR和游离的抗体分子。 (2)由连续的或不连续的氨基酸残基组成。后者系抗原分子折叠后形成,因而不连续性表位又称构象表位或构象决定簇。蛋白质的变性和抗原分子的分解将破坏构象,此时,能识别天然蛋白质的抗体即不再起作用。另外,组成不连续表位的肽链间如果由二硫键相连,则此键一旦被打断,表位即被破坏。 构象表位的形成和 ...
抗原类别的区分根据抗原是否显示免疫原性而区分抗原 (1)完全抗原:即免疫原。根据化学特性,免疫原性最强的是蛋白质抗原,多糖次之;脂类和核酸如果与蛋白质及多糖形成复合物,可显示免疫原性。 (2)半抗原:因分子质量小仅具有抗体结合特异性,需和载体蛋白连接后方具有免疫原性。此时,半抗原激活B细胞,载体蛋白激活T细胞。换言之,对于半抗原―载体系统,B细胞表位和T细胞表位分别位于半抗原和载体分 ...
受体分子启动的信号转导和基因的转录激活 所有受体分子和配体(抗原)相互配接后启动信号转导,舍此,免疫细胞不能激活,免疫应答也难以有效地进行。通过信号转导,细胞外信号被转换成胞内的生化事件,引起基因的转录激活和产物表达。 蛋白磷酸化、蛋白激酶和蛋白磷酸酶 蛋白质肽链某些氨基酸残基上的氢原子可以被ATP释放的磷酸根取代而发生磷酸化。如果因磷酸化而被修饰的蛋白质属于酶,可使其处于 ...
淋巴细胞的多克隆激活剂 多克隆激活剂可使高比例的淋巴细胞活化。严格说来,这类激活剂不属于抗原,因为它可激活大量T、B细胞克隆而不涉及这些克隆的抗原特异性。但是多克隆激活剂可引起强烈的免疫病理学效应,在免疫应答的正负调节及其体外应答和临床应用上,有重要价值。淋巴细胞多克隆激活剂 (1)丝裂原:能引起高百分比的T、B细胞发生有丝分裂的物质称为丝裂原(mitogen)或有丝分裂原。丝裂 ...

参与免疫细胞激活信号转导的蛋白磷酸酶 蛋白磷酸酶的作用和蛋白激酶相反。根据脱磷酸化的氨基酸残基的不同,蛋白磷酸酶也分成蛋白酪氨酸磷酸酶(PTP,PTPase)和丝氨酸/苏氨酸磷酸酶。参与淋巴细胞激活的蛋白磷酸酶主要有: ①CD45:该分子胞内段的两个结构域发挥PTP的作用,因而CD45属于受体型蛋白酪氨酸磷酸酶,在对抗瓢kPTK的作用和启动淋巴细胞信号转导中发挥关键作用; ...
免疫细胞的各种受体 免疫系统对抗原的识别主要通过受体实现。这一识别涉及受体分子的胞外部分和配体(抗原)的专一性配接(ligatlon),胞内部分(或是受体相关分子的胞内部分)则负责抗原识别的信号转导(slgnaltrahsdUCtlon),由此引起免疫细胞的激活。 1.抗原识别受体 所有的非己成分原则上都可以作为抗原被识别而启动免疫应答。因抗原不同,受体主要分成固有免疫中识别PAM ...
蛋白丝氨酸/苏氨酸激酶 功能是使蛋白质上的丝氨酸和苏氨酸残基发生磷酸化。除了参与协同刺激信号的启动之外,这类激酶一般在信号转导的中下游发挥作用。由于信号途径具有相互“串流”(cross-talking)的特点,蛋白丝/苏氨酸激酶往往为各种免疫细胞和不同受体介导的信号途径所共用。与免疫细胞活化有关的丝/苏氨酸激酶主要有: 蛋白激酶C(PKC):参与淋巴细胞激活信号转导的磷脂酰肌醇途径 ...
脂筏与信号途径 免疫细胞信号转导的通用简要流程中,受体跨膜分子附近的细胞膜形成一种特化性结构称为脂筏(1ipidraft)。这是一种能够抗去污剂作用,富含糖脂的微结构域(glycolipid -enrlchedmicrodomaln,GEM),由鞘脂(spingolipid)、胆固醇和糖基磷脂酰肌醇(GPI)锚定蛋白等组成,质地较硬,能够像船筏一样漂浮在周围流动性较强的膜脂双层结构中。脂筏 ...
转录因子和衔接蛋白 转录因子又称核因子(nuclear{actor)或反式作用蛋白,通常以未激活状态游离于胞质,一旦激活即进入细胞核,称为转位(translocation),与相应基因启动子区特定的DNA框(DNAbox)即顺式作用元件相结合,最终激发通用转录复合体(GTC),启动基因转录。转录因子的激活,是发生信号转导级联反应的结果,和上述PTK和PTP对转录因子的磷酸化和脱磷酸化有关。 ...
适应性免疫涉及的一些重要概念1.三个特点 (1)特异性:二次应答时能精细地区分致敏的抗原和其他无关的抗原;因而免疫应答的特异性指的是抗原特异性 (2)多样性:参与获得性免疫应答的淋巴细胞,其表面识别结构和相应分子(如抗体)所显示的高度异质性,赋予机体具有识别数量极大的抗原并与之起反应的能力。多样性是特异性产生的基础。 (3)记忆性:再次遇到同一抗原时,出现增强性应答。免疫记忆由记 ...
固有免疫和适应性免疫应答的相互联系 没有固有免疫细胞和分子的介入,就不会出现有效的特异性免疫应答。固有免疫和适应性免疫是免疫系统不可分割的两个方面。 1.参与固有免疫和适应性免疫的细胞和分子在结构和功能上的关联性 (1)免疫细胞:参与固有免疫(Me、NK细胞)和适应性免疫(T、B淋巴细胞)的细胞间存在兼有两者特性的免疫细胞,如前面提到的固有类淋巴细胞NKT、78T和B1细胞。 ...
关于丁香通
公司信息
个人用户
企业机构


