


二价体染色体的非姊妹染色单体间发生交换的形态表现,最早出现在双线期。交叉在各条染色体上并无特定部位和数目,但交叉点数目与重组节数目大体相等,而且随着时间的推移交叉点向末端移动。端化过程一直进行到中期Ⅰ,随着端化过程的发展,交叉点位置不断移动,交叉数目减少。
物质通过质膜进出细胞的方式之一。物质从高浓度一侧通过质膜向低浓度一侧的扩散,不需要载体也不消耗能量。物质的自由扩散速度,与物质的脂溶性程度、膜两侧溶质浓度差、溶质分子大小和电荷性质等有关。由于脂类双分子层构成质膜的基本骨架,脂溶性物质能够溶于膜脂内,因此能优先通过细胞膜进出细胞。大量实验表明,许多物质通过质膜的扩散速度都与其脂溶性程度成正比。水几乎不溶于脂,水分子可以经过质膜上的小孔自由进出细 ...
物质通过质膜进出细胞的方式之一。是质膜上载体蛋白消耗能量而协助物质逆浓度梯度运输的方式。主动运输和协助扩散虽然都是在载体蛋白协助下运输物质,但两者有两个主要区别:一是主动运输是质膜上载体蛋白逆浓度梯度运输物质;二是主动运输与能量代谢联系,载体蛋白消耗ATP提供的能量。例如,枪乌贼神经纤维内部Na 含量相当于外部Na 含量的十分之一。如果向枪乌贼的巨大神经纤维内部注射24Na ,不久即测得神经纤 ...
氨基酸用肽键相连构成的生物大分子,是细胞最重要的功能成分,约占原生质有机成分的一半。 功能 基因表达成蛋白质,才能体现生物的特性。蛋白质的功能多种多样,可列举如表。 分类 整个生物界的蛋白质有百万种以上,分类方法如下: 依据功能分类 除贮存蛋白质和结构蛋白质外皆为功能蛋白质(酶、调节蛋白、运动蛋白,运输蛋白、防御蛋白等)。 依据多肽链的数目分类 只含一条多肽链的蛋白质称单链蛋白质;含有 ...
减数分裂Ⅰ的偶线期中,配对的两条同源染色体之间形成的一种复合结构。它对于维持同源染色体配对的稳定性,以及同源染色体的局部交换,是不可缺少的条件。联会复合体是由核蛋白组成的扁带状三分区结构,总宽度约为150~200纳米。在减数分裂Ⅰ的细线期,每个染色体的两条染色单体之间出现一种宽约30纳米的线状结构,该结构沿染色体全长分布,其两端都与核膜相接触,由它发育成联会复合体的侧生组分。在偶线期中同源染色 ...
允许溶剂(水)分子通过,不允许溶质分子通过的膜。生物膜、火棉胶膜、玻璃纸等均非理想半透膜,它们在不同程度上允许溶质分子通过,但通过水分子的量比溶质分子多很多。人工方法制造的亚铁氰化铜的沉淀膜是接近于理想的半透性膜,它是在硫酸铜溶液与亚铁氰化钾溶液的接触面上形成的:2CuSO4 K4Fe(CN)6→Cu2Fe(CN)6 2K2SO4。该沉淀膜很纤弱,不能单6独使用,在制做时必需有支撑物。 ...
细胞内的分子排出或挤出到周围介质中。分泌物包括离子、酶、激素、糖蛋白、液体、无机盐等。分泌的主要形式如下。 局部分泌(merocrine) 或称外分泌(exocrine),为最普遍的胞吐方式。其分泌物为小的包以膜的颗粒与质膜融合后而排出的复杂过程。如消化酶原颗粒的分泌过程包括:分泌物所需物质从微血管运送至腺细胞基部线粒体之间;线粒体为三磷酸腺苷生成提供能量,三磷酸腺苷获取的能量用于合成有机物 ...
原核生物的一个类群。大多数有发达的分枝菌丝。菌丝纤细,宽度近于杆状细菌,约0.5~1微米。可分为:营养菌丝,又称基质菌丝,主要功能是吸收营养物质,有的可产生不同的色素,是菌种鉴定的重要依据;气生菌丝,叠生于营养菌丝上,又称二级菌丝。在气生菌丝上分化出可产生孢子的孢子丝,孢子丝的形状和排列方式因种而异。成熟的孢子丝上产生成串的分生孢子。孢子的表面结构、形状及颜色在一定条件下比较稳定,是鉴定菌种 ...
蛋白质生物合成过程。生物都有从氨基酸合成自身蛋白质的能力,此过程在核糖体上进行,信使RNA(mRNA)是合成的模板。生物依照mRNA的密码子序列,通过转移RNA(tRNA)的反密码子与密码子配对,使相应的氨基酸从N端到C端依次参入蛋白质。这个过程十分复杂,有几百种生物分子参与。以原核细胞为例,大体可分为两个阶段: 氨酰tRNA合成酶识别特定的氨基酸及对应于该氨基酸的tRNA,并催化氨酰tRN ...
缝隙连接分布广泛,几乎存在于所有的动物细胞中。用超薄切片术可显示相邻两细胞的连接处的细胞质膜明暗相间七层结构,细胞间的缝隙约2纳米,其内有间隔的均匀排列的颗粒。用冰冻断裂电镜技术显示缝隙连接的颗粒区面积大小不等,且排列规则而密集。用X线衍射技术证明,每个颗粒由6个蛋白质亚单位构成,它们呈环行排列,中间有直径2纳米左右的小孔,被称为连接小体(con-nexon)。每两个连接小体相对合,并分别包 ...
大量微管纵向排列组成的中间宽两极小的细胞器,形状象纺锤,因而得名。因为纺锤体和染色体运动密切相关,所以纺锤体是一个重要的有丝分裂装置。纺锤体由极间丝(又叫连续丝或称极间微管)、着丝点丝(又叫染色体牵丝)、星体丝及区间丝四种微管组成。极间丝,指一极与另一极相连的纺锤丝,但绝大多数极间丝(连续丝)并非真正连续,而是来自两极的微管在赤道面彼此相搭,侧面结合。有的微管和两极均不接触。着丝点丝是指一端由 ...
亦称高尔基复合体、高尔基器。是真核细胞中内膜系统的组成之一。为意大利细胞学家高尔基Golgi于1898年首次用银染方法在神经细胞中发现。是由光面膜组成的囊泡系统,它由扁平膜囊(saccules)、大囊泡(vacuoles)、小囊泡(vesicles)三个基本成分组成。扁平膜囊是一扁平囊状结构,囊腔中央较窄,周边较宽,它们平行排列类似扁盘堆叠结构,形成扁平膜囊堆,亦称高尔基堆(Golgi st ...
真核细胞核内无界膜包围的球状小体。通常有1~2个,也有多个。核仁大小与数目因生物种类的生理状态不同而异。光镜下观察核仁为匀质球体,电镜观察其亚显微结构,有三个特征性区域:(1)颗粒区(granular component),内含电子密度较大的颗粒,直径为150~200埃(),是处于不同成熟阶段的核糖体亚单位的前体;(2)纤维区(fibrillar component),由直径50~100埃的 ...
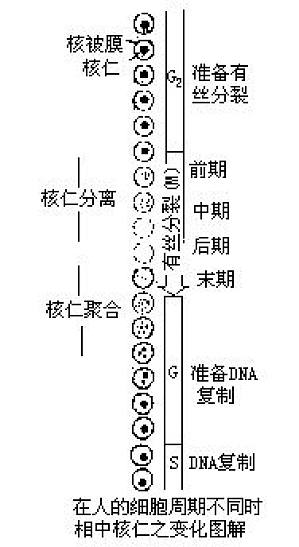
在细胞周期中,核仁进行分离和重新聚合的过程。当细胞进入有丝分裂时,核仁首先变形和变小;其后染色质凝集和停止核糖核酸(RNA)合成,包含有核糖体RNA(rRNA)基因的DNA袢环逐渐收缩回到相应染色体的核仁组织区;核膜破裂进入中期,这时核仁消失;在有丝分裂末期时,核仁组织区DNA解凝集,rRNA合成重新开始,极小的核仁重新出现在染色体核仁组织区附近。核仁形成后常发生融合现象。如人二倍体细胞在相 ...
参与形成核仁时的染色质区,核仁从核仁组织区部位产生,同时与该区紧密相连。具有核仁组织区的染色体称核仁染色体。核仁组织区定位在核仁染色体次缢痕部位。对人来说,在13、14、15、21、22对染色体上存在核仁组织区。核仁形成后常发生融合现象,所以虽存在多对染色体,但常见间期细胞中仅有1~2个核仁,如人二倍体间期细胞中含一个大的核仁,它包含有从5对核仁染色体上核仁组织区来的DNA袢环,这些袢环上含有 ...
真核生物染色质包装的基本结构单位。染色质纤维之基本结构是由核小体串连而成。它通常含有200个碱基对的脱氧核糖核酸(DNA)和9个组蛋白分子。由核小体核心(nucleosomecore)和一条含有H1组蛋白的连接区DNA(linkerDNA)所组成。核小体核心由四种组蛋白(H2A、H2B、H3、H4各2个分子)小体核心结合上H1组蛋白后,165个碱基对的DNA分子在组蛋白八聚体上缠绕2圈。H1组 ...
亦称核蛋白体、核糖核蛋白体。是普遍存在于各类细胞中的无被膜的颗粒结构,为细胞合成蛋白质的重要场所。除存在于细胞质(游离态)、线粒体和叶绿体中外,也结合排列在内质网膜和核膜外表面上,后者主要合成向细胞外输送的分泌蛋白和装配内膜系统之蛋白。核糖体直径约15~30毫微米,包含大、小两个亚单位,大、小两亚单位间有一个被称为隧道的间隙,其中有信使核糖核酸(mRNA)细丝通过。原核细胞核糖体沉降系数为70 ...
核糖体的主要组分,可达其重量的65%,原核生物中有3类rRNA,而真核生物核糖体含有4种rRNA。rRNA在构成核糖体结构和行使其生物合成功能上均有重要作用。 ...
催化RNA合成的酶。主要指DNA指导的RNA聚合酶,即以DNA为模板,以4种核苷三磷酸(ATP,GTP,CTP和UTP)为底物,从5′到3′合成RNA的酶。其催化方式与DNA聚合酶相似,但不具备有校正作用的外切核酸酶活性,聚合反应也不需引物。 对大肠杆菌RNA聚合酶了解得最多。这种酶含有多个亚基,全酶结构为α2ββ′σ。 σ亚基易从全酶上脱落下来,剩下的α2ββ′部分称核心酶,有催化活性。 ...
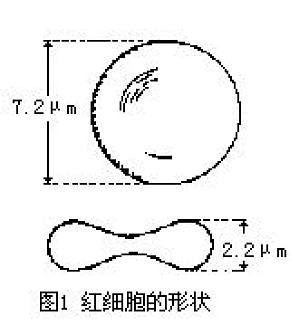
血液中最多的一种血细胞,红细胞总数,我国正常成人,男约450~550万个/毫米3,女约380~460万个/毫米3。人和哺乳动物的成熟红细胞无细胞核,含血红蛋白。直径约8微米(μm),呈双凹圆碟形,中央部分较薄,边缘部分较厚,平均厚度约2微米,表面积约120微米2。此细胞形状的表面积与体积之比值增大(与球形相比),有利于气体扩散出入红细胞,也有利于可塑性变形,当红细胞挤过口径比它小的毛细血管和血 ...
关于丁香通
公司信息
个人用户
企业机构


