大家都在搜


对模式生物秀丽隐杆线虫(C. elegans)而言,在其基因组特定位点引入单拷贝外源基因是一种常见的研究模式。ttTi5605基因座表达载体是一种经过良好验证的可用于线虫的特定位点的外源基因单拷贝插入的高效系统。该系统使用Mos1转座子剪切机制或CRISPR机制向线虫引入稳定整合的外源基因插入。
Mos1转座子
Mos1转座子首先在毛里求斯果蝇(Drosophila mauritiana)中被发现。当前,相当一部分的线虫种属已经被开发出包含确定的Mos1转座子插入位点。一个例子是基因组ttTi5605区域包含了Mos1位点的线虫种属。使用ttTi5605基因座的原因在于在该区域插入序列不会扰乱临近基因的功能。此外,Mos1转座子剪切技术可以将外源基因以单拷贝的方式插入基因组,不容易被线虫自身的RNAi系统沉默(重复元件序列通常易被细胞转录抑制)。
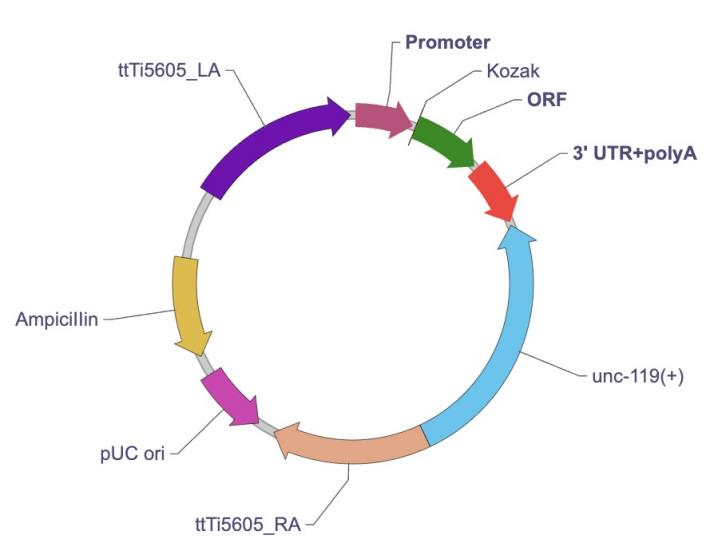
Mos1介导的ttTi5605位点的基因特异性重组需要ttTi5605基因座表达载体与一个表达转座酶的辅助质粒共转染线虫。通过热激法可以激活转座酶活性,从而导致Mos1转座子在线虫基因组发生剪切。断开的DNA双链可以被外部提供的序列修复并产生稳定的基因重组。通过Mos1介导的单拷贝插入(Mos1-mediated Single Copy Insertion,MosSCI)技术,目的基因可以被整合至线虫的ttTi5605位点。 ttTi5605基因座表达载体应用 ttTi5605基因座表达载体包含一系列重要元件帮助高效表达目的基因。 最后,载体还引入了一个野生型C. Briggsae线虫的unc-119基因作为阳性筛选标记。Unc-119(ed3)突变的线虫体型小、呈现出运动不协调表型,并且在缺乏食物时不能进入dauer状态。转染ttTi5605基因座表达载体后,只有恢复野生型表型的线虫表明转染是成功的。
ttTi5605基因座表达载体图谱
使用Mos1转座子产生的DNA双链重组是稀少事件,而更大范围的基因组修饰可以使用CRISPR技术。ttTi5605基因座表达载体也能应用于Cas9触发的DNA同源重组。针对这种应用,一个打靶ttTi5605位点临近序列的sgRNA载体与一个Cas9蛋白需要被转染至线虫体内。ttTi5605基因座表达载体可以将外源基因高效插入线虫基因组。连同表达转座酶的shRNA/Cas9载体,ttTi5605 Mos1靶向质粒可以实现位点特异的单拷贝外源基因插入。 ttTi5605基因座表达载体的特点 ttTi5605基因座表达载体可以将外源基因高效插入线虫基因组。连同表达转座酶的shRNA/Cas9载体,ttTi5605 Mos1靶向质粒可以实现位点特异的单拷贝外源基因插入。该载体从机制和使用上拥有两大优势: 1.载体DNA持久整合。向线虫转染普通质粒只能实现瞬时转染,外源基因将随时间在宿主细胞内逐渐丢失。而ttTi5605基因座表达载体可以整合转基因至细胞基因组并稳定表达,可用于基因序列的精确编辑以及构建转基因线虫。 2.操作技术简单。向线虫转染质粒载体是技术上相对直接且对比病毒载体需要包装生产而更为简单的方式。可以使用电转染、显微注射或者喂食包含质粒载体的大肠杆菌等方式将质粒导入线虫体内。 ttTi5605基因座表达载体也有一定的不足之处,比如说MosCI系统不能同时编辑多个基因。此外,使用该系统获得转基因动物模型通常需要超过15天。 更多线虫ttTi5605基因座表达载体的信息,可后台或电话(020-28069042)咨询
【参考文献】
1. Nature. 413:70 (2001) 2. Nat Genet. 40: 1375 (2008) 3. Worm. 4:e1046031 (2015) 4. Curr Biol. 1476:82 (2008) |

更有优质直播、研选好物、福利活动等你来!