大家都在搜

| 简介 由于许多技术的进步和“多组学”的应用,表观遗传学已从基因组区域的单一表观遗传控制的简单解析层面向更深层次的方向发展。 我们现在可以结合转录组学分析和相关分析平台分析全基因组组蛋白修饰模式、转录因子结合谱、染色质可及性谱、三维染色体构象和DNA甲基化动力学。 一系列引人入胜的研究已经开始向我们展示如何在不同的表观遗传学水平上整合多种NGS技术,如何支持构建多因素的表观转录图谱。 这些研究工具可以揭示控制基因表达的潜在机制,以及它们是如何影响正常或病理生物学过程。
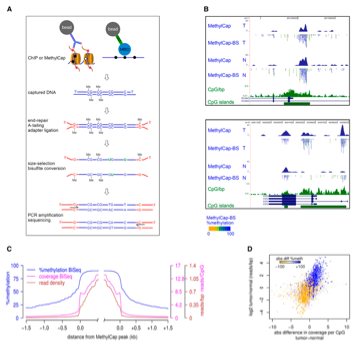
本文重点介绍了一系列利用基于多组学分析的研究,研究人员将ATAC-Seq、RNA-Seq、DNA甲基化和MeDIP-Seq以及ChIP-Seq等方法结合起来,对多种类型的细胞和组织进行分析。 这些研究都采用多组学方法揭示了正常发育、疾病和癌症的表观遗传学过程。 心肌细胞发育——触及心脏的核心 在心血管发育模型中的多组学分析中,Kranzhofer等人试图深入了解新生和成年小鼠心肌细胞(负责在心脏中产生收缩力的细胞)的表观遗传学动力学,以理解为什么这些至关重要的细胞在出生不久后细胞成熟时会失去增殖能力。 使用5-羟甲基胞嘧啶测序(hMeDIP-Seq)、5-甲基胞嘧啶测序(MeDIP-Seq)、RNA-Seq,以及利用新生和成年小鼠中分离的心肌细胞做染色质免疫共沉淀(ChIP)-Seq(H3K27ac),将其结果结合分析,作者证明了5-hmC修饰与随后表现出上调表达的基因(Mb和Pdk4基因)的基因体中DNA甲基化缺失正相关。 总的来说,这项基于多组学的研究有助于更好地定义小鼠心肌细胞成熟过程中基因表达的表观遗传学控制,这可能有助于未来的治疗性表观遗传学干预,即“启动”非增殖性心肌细胞以修复人类患者心肌梗死后受损的组织。[1] 胚胎干细胞——影响细胞命运的相互依赖关系 在这项工作中,Brinkman等人利用ChIP-bisulfite-seq(图一)定量评估了与组蛋白修饰H3K27me3相关的小鼠胚胎干细胞(来自早期植入前胚胎内部细胞团的多能干细胞)的DNA甲基化模式。 研究基因组的表观遗传修饰有助于作者理解DNA甲基化和H3K27me3之间的相互依赖性。 令人感叹的是,这项研究揭示了除了CpG岛(通常形成相关基因调控区的DNA甲基化敏感区)外,H3K27me3和DNA甲基化在整个基因组中共存,其中两个修饰显示出相互排斥性(与Plekha2/Htra4和Lmx1b/C130021I20Rik基因相关的CpG岛)。 这项研究者认为,类似的研究可以提供更多关于不同染色质形式的复杂组成及其在胚胎干细胞中的生物学作用的见解,从而支持高效产生治疗相关细胞类型的高级分化方案。[2]
图一: 捕获方法与亚硫酸氢盐深度测序的整合: ChIP-BS-seq和MethyCap-BS-seq 基于多组学的表观遗传学分析——植物中的多组学 zhong等人的一项研究试图利用多组学技术来了解DNA甲基化如何影响染色质可及性、高阶基因组组织和基因表达。 作者们试图在模式植物拟南芥中探索这种迷人的联系。作者将DNA甲基化数据与染色质可及性分析(通过使用ATAC-Seq分析染色质可及性)和高阶染色体构象分析(通过高通量染色体构象捕获Hi-C测序)(图二)相结合。 这项研究的发现包括,染色质可及性的增加并不总是导致基因转录的增加,以及DNA甲基化可能通过其他机制影响染色质结构的结论。 总之,作者们发现了特定DNA甲基化模式、染色质可及性和三维基因组结构之间紧密的联系,并揭示了DNA甲基化直接影响这个重要模型系统中的染色质结构。[3]
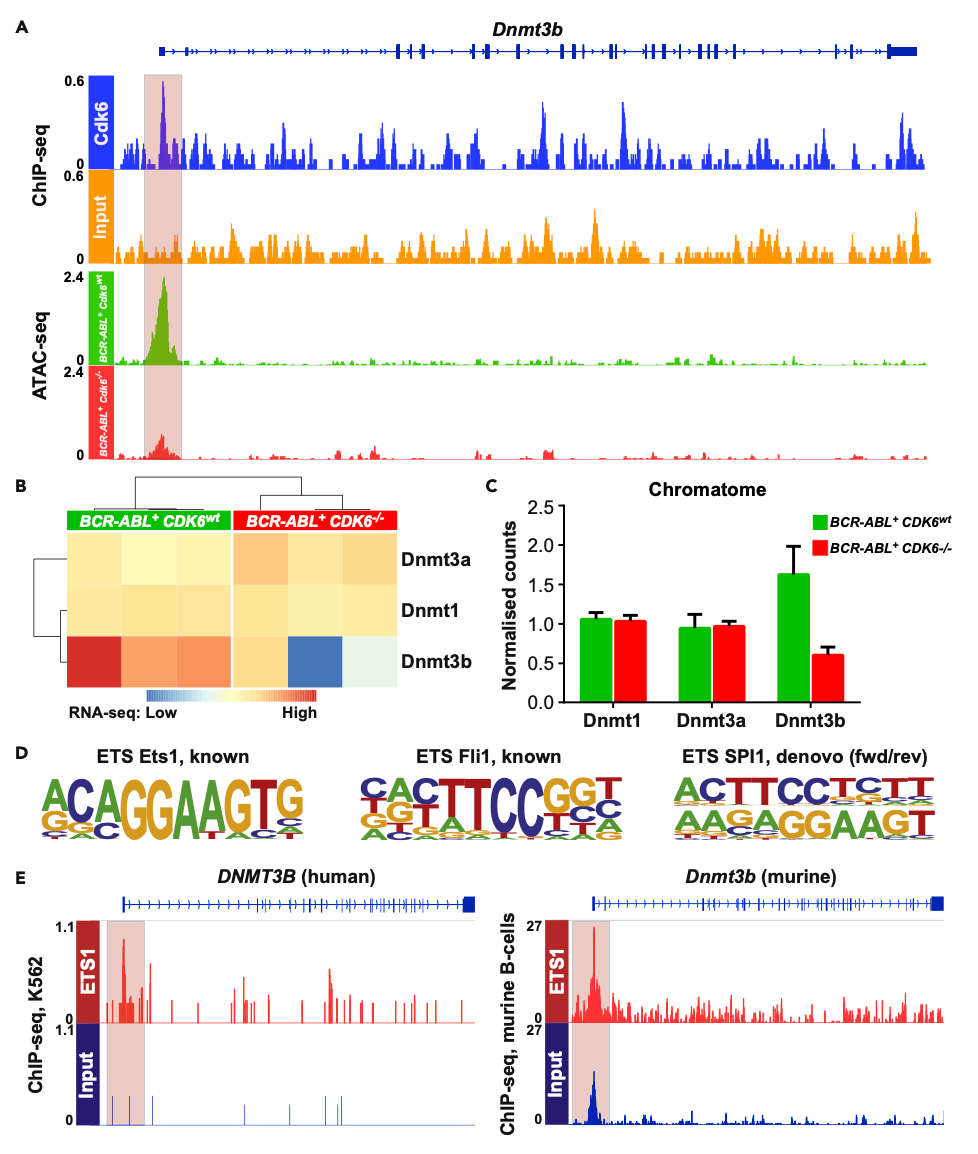
图二: met1中的染色体构象信号再分配 疾病的多组学分析——揭示癌症的新机制和治疗目标 Heller等人将DNA甲基化分析与ChIP-Seq、ATAC-Seq和RNA-Seq相结合,以描述急性淋巴细胞白血病(ALL)患者细胞中一种关键调节机制的失调——CDK6介导的CpG岛DNA甲基化调节。 他们基于多组学的方法强调了CDK6(在许多癌症类型中过度表达)如何控制多个水平的调控,包括经典基因调控和表观遗传调控。 值得关注的是,本研究整合了CDK6的ATAC-Seq和ChIP-Seq数据,以展示CDK6如何与Dntm3b基因结合,从而增加染色质可及性,促进基因表达,从而诱导全基因组DNA甲基化(图三);同时,诱导的CDK6表达缺失降低染色质可及性,抑制Dnmt3b表达,并降低DNA甲基化水平。 总之,CDK6作为癌细胞DNA甲基化调节因子的发现,支持了CDK6活性小分子修饰剂作为潜在有趣的抗ALL疗法的探索。[4]
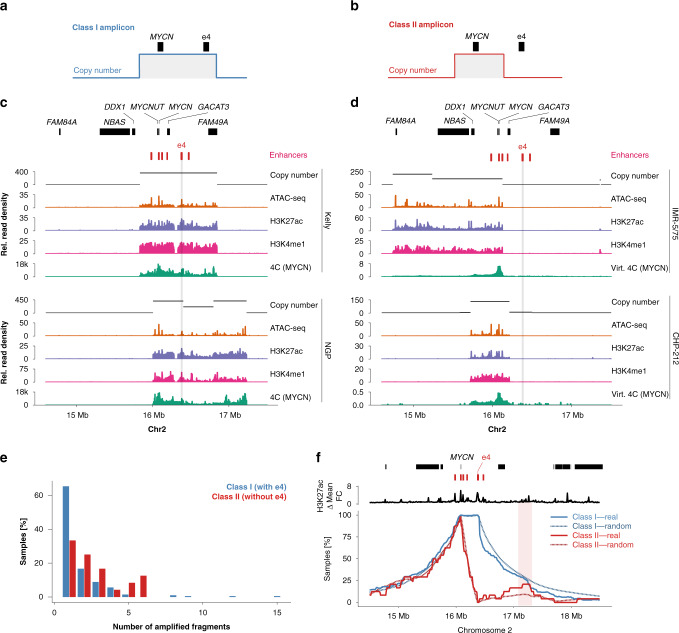
图三:DNMT3B在BCR-ABL+细胞中受CDK6转录调控 多组学在神经母细胞瘤中的应用 神经母细胞瘤是一种在未成熟神经细胞中发生的癌症,每6例中就有1例出现MYCN基因的扩增,在高度重排的染色体外环状DNA颗粒上通常会发现额外的基因拷贝。 Helmsauer等人通过ChIP-seq对神经母细胞瘤细胞系中的H3K27ac和H3K4me1进行了分析,ATAC seq和环状染色体构象捕获(4C)进行了基于多组学的MYCN基因座分析(图四),并将这些发现与来自短读长和纳米孔测序的MYCN扩增子结构相关数据相结合。 总之,这项激动人心的研究报告称,“异位增强子劫持(ectopic enhancer hijacking)”驱动了MYCN的表达,这有助于解释观察到的结构多样性以及与MYCN扩增相关的表观遗传调控。[5]
图四: 在神经母细胞瘤中可以识别出两类MYCN扩增子 血癌——多方助力血癌研究 基于多组学的疾病评估的最后一个例子涉及多篇关于慢性淋巴细胞白血病(CLL)的研究论文(Beekman等人、Ott等人,和Rendiero等人),这些论文构成了Active Motif(“血癌研究的表观遗传学工具”)最近一次网络研讨会的基础。 这些基于CLL的研究希望探索与疾病相关的表观基因组和调控网络的改变,确定遗传和表观遗传结构之间的关系。 他们的工作旨在通过了解疾病异质性的基础来确定潜在的治疗目标。 作为研究的一部分,Beekman等人评估了原代CLL和正常细胞中与ATAC-Seq、DNA甲基化谱和RNA-Seq相关的CLL基因座(FMOD和TCF4基因座,原文图3)[6]。 类似地,Ott等人利用H3K27ac ChIP-seq、ATAC-seq和转录因子结合位点分析来了解CLL细胞中超级增强子位点的疾病相关动力学(原文图1-3)[7]。 最后,Rendiero等人对CLL细胞中H3K4me1、H3K27ac和H3K27me3的RNA-Seq、ATAC-Seq和ChIPmentation(使用Tn5介导的标记制备ChIP-seq测序文库)分析了CLL相关基因座(Rendiero等人;ZNF667和ZBTB20基因座,原文图3F)[8]。 总之,这些研究大大有助于我们理解与疾病相关的表观遗传学机制,有助于识别CLL亚型,并为成人最常见的白血病的表观遗传学治疗提供了基础,其中包括可能应用表观遗传学“readers” BET蛋白家族抑制剂。 单细胞多组分析——建立更大的组学数据库 虽然上面讨论的表观遗传学技术通常使用数百万个细胞作为输入,并使用不同的细胞样本,整合来自不同实验的数据集,但某些细胞样本(包括患者样本)的稀缺性是基于多组学的评估的一个重大障碍。 Active Motif现在推出了一项名为“单细胞多组”的新服务,提供了一种解决方案。这项技术由10 × Genomics开发,将单细胞(sc)ATAC-seq与scRNA-seq相结合。 Swanson等人提供了这种方法潜力的一个例子,作者创建了一个基于液滴的多组学平台,支持数千个单细胞中同时scRNA-Seq、scATAC-Seq和蛋白质丰度检测,他们称之为TEA-Seq(transcriptomics、epitopes和accessibility)。 作者希望类似的多模式单细胞分析能够支持在表型定义罕见的细胞类型群体中鉴定细胞类型和其特异性基因的表达,以及调控[9]。 单细胞多组学可以消除需要分选策略的方法,如FACS或磁分选,这些方法可能会由于样本处理本身而改变细胞的生物学特性,而单细胞多组学可以真正了解细胞的真实表观遗传特征。
此外,对于表观遗传研究中常用的实验技术——包括ChIP-Seq、ATAC-seq、RNA-Seq、DNA甲基化分析等,Active Motif有超过20年的相关技术服务经验: 我们提供○ChIP-Seq服务 ○CUT&Tag服务 ○ATAC-Seq服务 ○单细胞ATAC-Seq服务 ○Mod Spec® 服务 ○RIME ○RNA-Seq服务 ○单细胞RNA-Seq服务 ○ChIP抗体验证服务 ○ChIP-qPCR服务 ○DNA甲基化服务 ○Hi-C服务
只需要准备好样本, 剩下的交给我们!
参考文献 [1] Kranzhöfer D K, Ralf G, BA Grüning, et al. 5'-Hydroxymethylcytosine Precedes Loss of CpG Methylation in Enhancers and Genes Undergoing Activation in Cardiomyocyte Maturation[J]. Plos One, 2016, 11(11): e0166575. [2] Brinkman A B, Gu H, Bartels S, et al. Sequential ChIP-bisulfite sequencing enables direct genome-scale investigation of chromatin and DNA methylation cross-talk[J]. Genome Research, 2012, 22(6):1128-1138. [3] Zhong Z, Feng S, Duttke S H, et al. DNA methylation-linked chromatin accessibility affects genomic architecture in Arabidopsis[J]. Proceedings of the National Academy of Sciences of the United States of America, 118(5): e2023347118. [4] Heller G, Nebenfuehr S, Bellutti F, et al. The Effect of CDK6 Expression on DNA Methylation and DNMT3B Regulation[J]. iScience, 2020, 23(10):101602. [5] Helmsauer K, Valieva M E, Ali S, et al. Enhancer hijacking determines extrachromosomal circular MYCN amplicon architecture in neuroblastoma[J]. Nature Communications, 2020, 11(5823). [6] Beekman R, Chapaprieta V, Russiñol N, et al. The reference epigenome and regulatory chromatin landscape of chronic lymphocytic leukemia[J]. Nature Medicine, 2018, 24(6). [7] Ott C J, Federation A J, Schwartz L S, et al. Enhancer Architecture and Essential Core Regulatory Circuitry of Chronic Lymphocytic Leukemia[J]. Cancer Cell, 2018. [8] Rendeiro A F, Schmidl C, Strefford J C, et al. Chromatin accessibility maps of chronic lymphocytic leukaemia identify subtype-specific epigenome signatures and transcription regulatory networks[J]. Nature Communications, 2016. [9] Swanson E, Lord C, Reading J, et al. Simultaneous trimodal single-cell measurement of transcripts, epitopes, and chromatin accessibility using TEA-seq[J]. eLife, 10: e63632.
原文作者:Stuart P. Atkinson, Ph.D |

更有优质直播、研选好物、福利活动等你来!