
大家都在搜

Cre-LoxP 系统是一种在 P1 噬菌体中发现的重组酶系统,通过利用重组酶 Cre 特异性识别 LoxP 重组位点的机制,可以在靶细胞和靶组织实现基因敲除、倒置和易位等基因编辑功能。在 P1 噬菌体中,Cre 重组酶识别 LoxP 位点其实是 X 基因侧翼的两个长度为 34 bp 的序列,由两个 13 bp 的反向重复序列和一个非对称的 8 bp 间隔序列组成。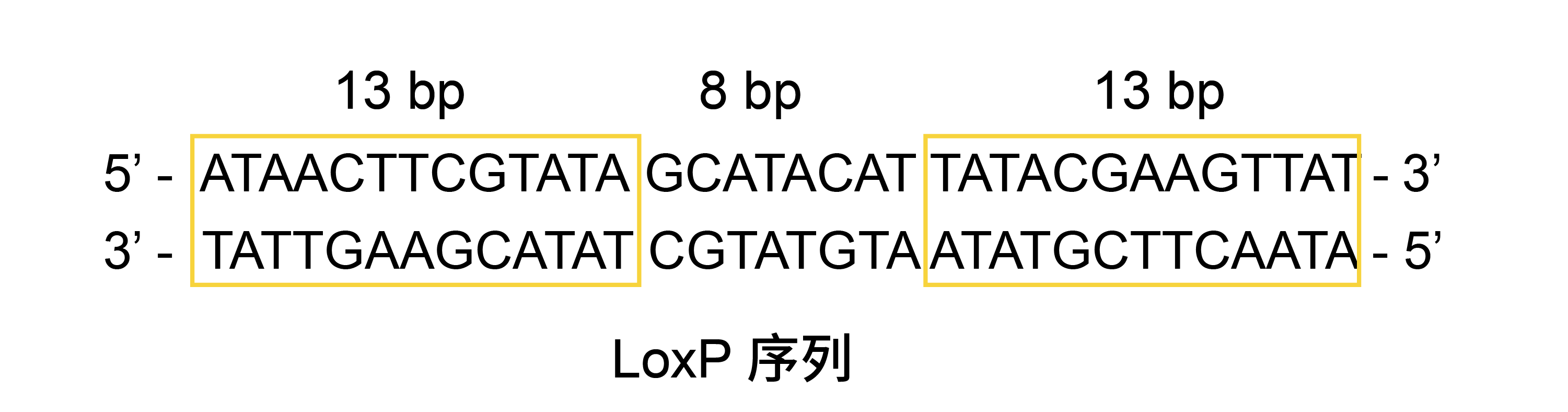 对于每个 LoxP 位点,Cre 重组酶的 2 个亚基高效地进行结合并形成二聚体 [2-5],然后该二聚体将与另一个 LoxP 位点上的 Cre 重组酶亚基二聚体形成联会复合物(Synaptic complex),催化 DNA 重组反应 [1,6,7]  由于 LoxP 位点的间隔序列是非对称的,LoxP 位点具有方向性。Cre 重组酶进行 DNA 重组时根据 LoxP 的方向,会产生不同的重组结果。 1. 如果一段序列上的两个 LoxP 位点反向,则重组后两个 LoxP 位点之间的序列被反转。 2. 如果一段序列上的两个 LoxP 位点同向,则两个 LoxP 位点之间的序列其中一个 LoxP 位点在 Cre 重组酶作用下被剪切出来。 3. 如果两条序列上具有同向的 LoxP 位点,则有可能发生两端序列的重组反应。 
重组反应是可逆的,但是对于剪切过程,由于切除出来的 DNA 序列为游离形式,重新发生整合的几率很小,因此 Cre-LoxP 系统可以有效被应用在基因敲除上。 我们通过将 Cre-LoxP 系统的原理从基因组编辑转换到质粒载体的序列编辑,构建出不同形式的含有 LoxP 位点的基因表达载体,则可实现 Cre 重组酶作用下该质粒载体的灵活的表达调控效果。 LSL 条件性表达系统 基于 Cre- LoxP 的基因调控表达系统的一种最基础方式即使用 LoxP-Stop-LoxP(LSL)表达盒。以 LSL 条件性表达质粒载体为例,LSL 表达盒位于目的基因上游和启动子下游之间,包含一个 3x SV40 polyA 以及其侧翼的同向 LoxP 位点。Cre 重组酶不存在的情况下,该表达盒阻断目的基因的正常表达。当引入 Cre 重组酶后,目的基因上游的 3x SV40 pA 被切除,从而目的基因能够在用户选择的启动子下被激活表达。 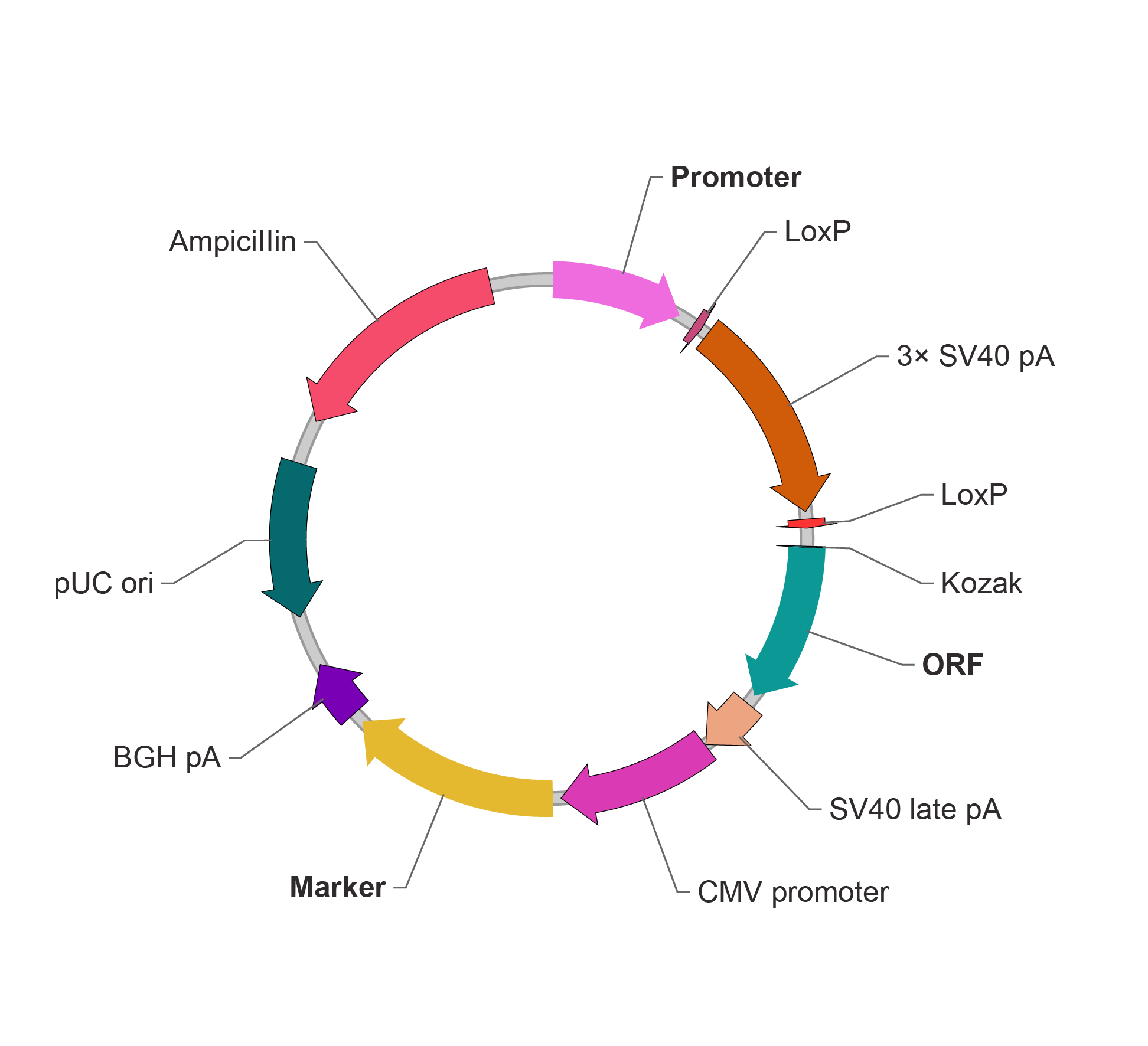
哺乳动物基因 LSL 条件性表达载体
FLEX 条件性表达系统 Flip-excision(FLEX)系统是另一种常用的基于 Cre- LoxP 的表达调控工具,该系统在一些文献中也经常被写作 Double-floxed inverse open reading frame,即 DIO 系统。FLEX 系统被广泛应用于要求时空特异性表达的神经示踪和光遗传学研究。 与 LSL 系统不同的是,FLEX 系统多增加了一对 LoxP 位点:Lox2272。Lox2272 是 LoxP 的突变体,但是不影响 Cre 重组酶的识别。只有同类的 LoxP 位点之间才能进行重组 [8],因此通过安排两种 Lox 位点的方向和位置,可以不可逆地改变目的基因的方向来实现其条件性表达。 具体设计原理如下 [9], 1. 默认状况下,目的基因的两翼设置交错且相向放置两个 LoxP 和两个 Lox2272。目的基因为反向,不表达。 2. 当引入 Cre 重组酶后,两个 LoxP 或者两个 Lox2272 之间的序列发生反转。 3. 重组产物中的同向 LoxP 或 Lox2272 位点进一步发生自我切除,使得目的基因两端最后形成不同类的 Lox 位点,因此不能进一步发生重组。 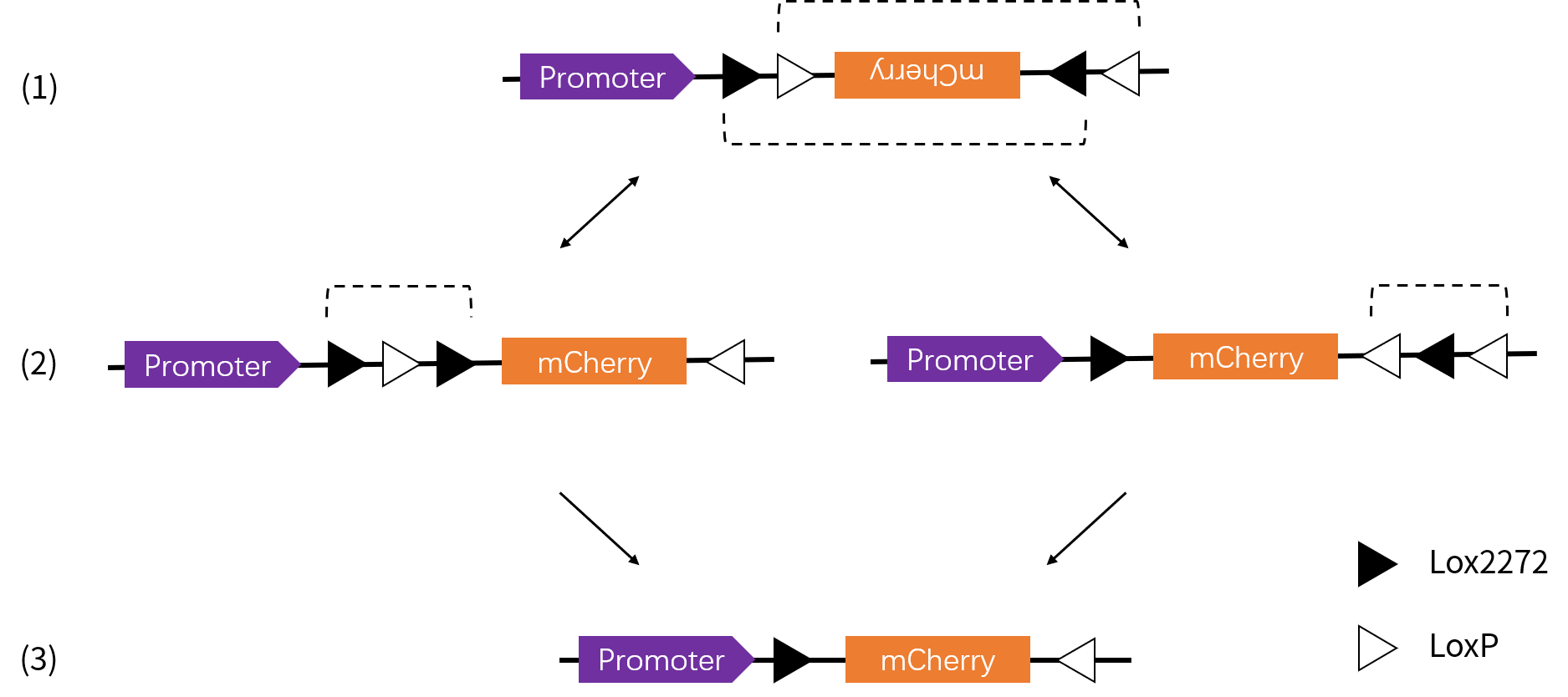 根据以上原理,可以进一步构造 ORF 默认方向不同的 FLEX 载体系统,即 Cre-On、Cre-Off 和 Cre-Switch。在 Cre 重组酶作用下,效果对比如下表:  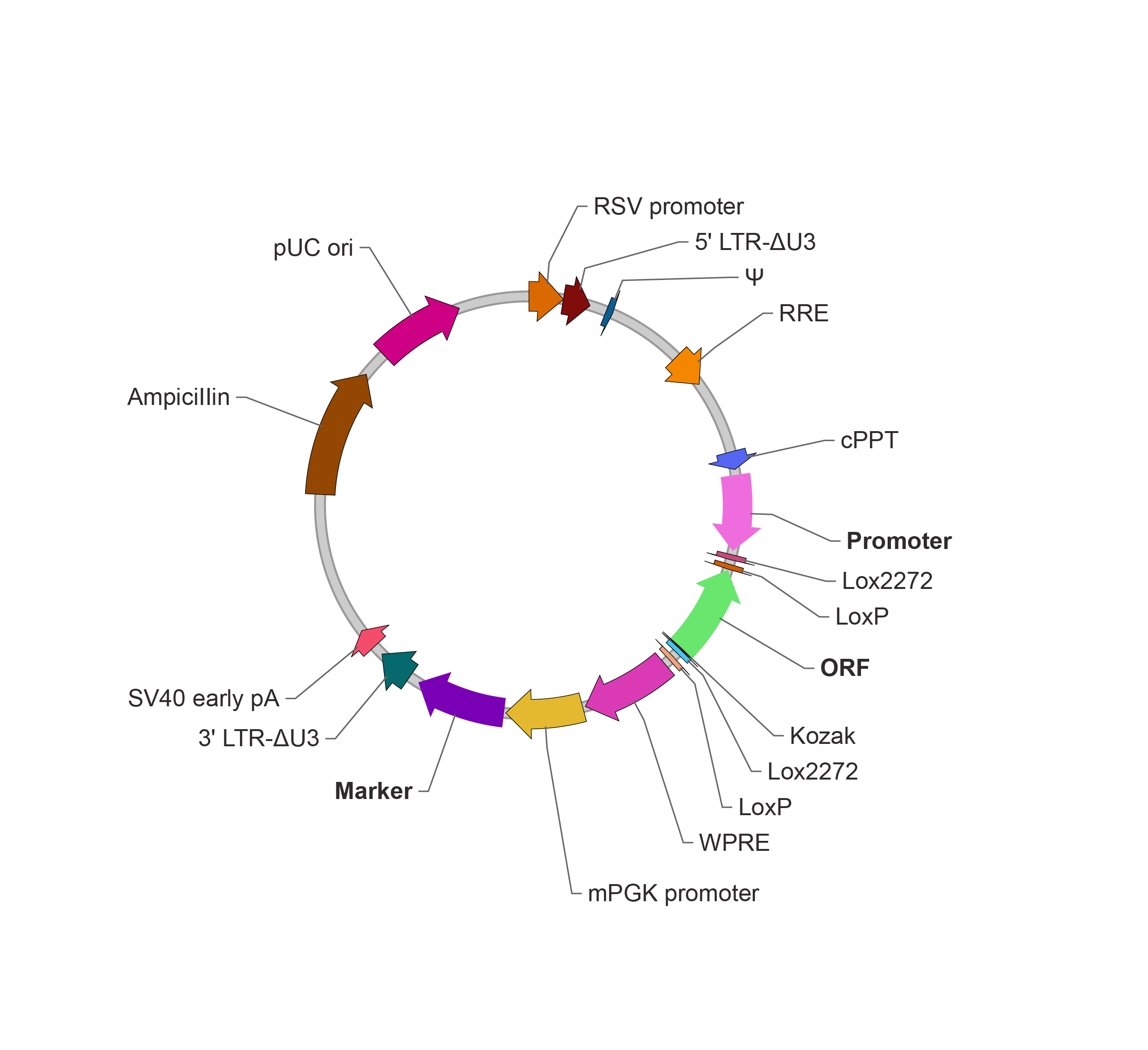
FLEX 条件性表达慢病毒载体(Cre-On)
ORF 默认与 Promoter 方向相反,Cre 重组酶介导的重组切割使 ORF 反转为与 Promoter 同向,ORF 表达。 
FLEX 条件性表达慢病毒载体(Cre-Off)
ORF 默认与 Promoter 方向相同,Cre 重组酶介导的重组切割使 ORF 反转为与 Promoter 反向,关闭 ORF 表达。 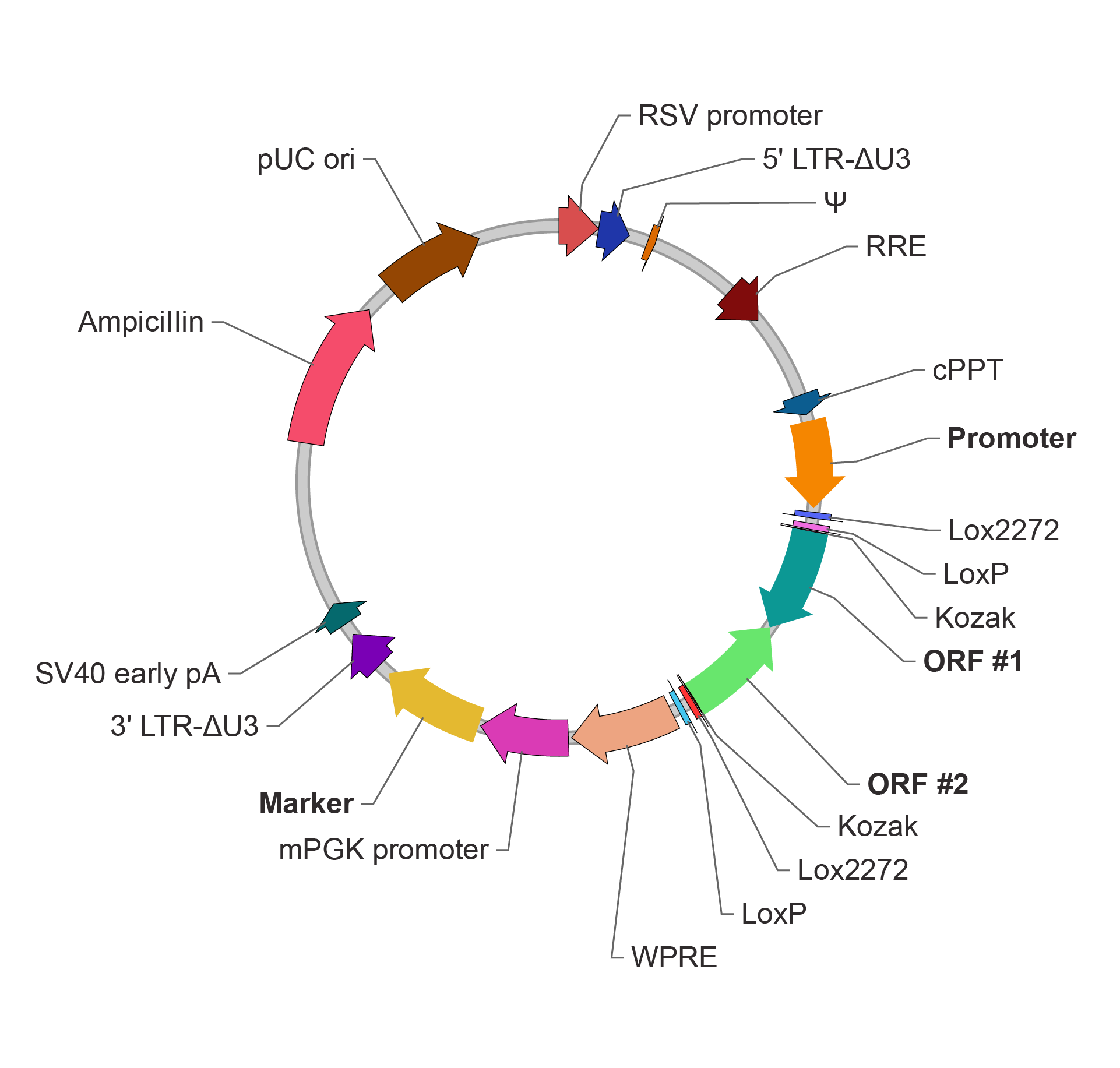
FLEX 条件性表达慢病毒载体(Cre-Switch)
ORF #1 默认与 Promoter 方向相同,ORF #2 则相反,Cre 重组酶介导的重组切割使 ORF #1 反转为与 Promoter 反向,关闭 ORF #1 表达。同时 ORF #2 也发生反转成为与 Promoter 同向,ORF #2 表达。诱导型 Cre-loxP 系统的关键:CreERT2 在一些情况下,我们希望在具有调控 ORF 表达的能力的同时,也需要精确调控 Cre 重组酶开始发挥作用的时间点。为此,利用诱导物诱导激活 Cre 重组酶因此也是常用的一种实验设计。CreERT2 是目前最常用的人工改造后的诱导型 Cre 重组酶,该酶融合了突变型的雌激素受体(Estrogen receptor,ER)的 LBD 结构域。默认状态下,CreERT2 通过其 ER 的结构域锚定在细胞膜内侧,此时不会进入细胞核编辑含有 loxP 的靶序列。添加 Tamoxifen 后,Tamoxifen 的代谢产物 4-Hydroxytamoxifen 则可以与 Cre-ERT2 结合,帮助其转运至核内从而发挥 Cre 重组酶作用 [10]。在表达 CreERT2 的工具鼠模型中,Tamoxifen 通过腹腔注射或者饮水给药均能发挥其诱导作用。 
CreERT2 表达慢病毒载体
参考文献
[1] Ghosh K, Guo F, Van Duyne GD. 2007. Synapsis of loxP sites by Cre recombinase. J Biol Chem. 282:24004–24016.
[2] Hoess R, Abremski K. 1984. Interaction of the bacteriophage P1 recombinase Cre with the recombining site loxP. Proc Natl Acad Sci USA. 81:1026–1029. [3] Mack A, Sauer B, Abremski K, Hoess R. 1992. Stoichiometry of the Cre recombinase bound to the lox recombining site. Nucleic Acids Res. 20:4451–4455. [4] Ringrose L, Lounnas V, Ehrlich L, Buchholz F, Wade R, Stewart A. 1998. Comparative kinetic analysis of FLP and cre recombinases: mathematical models for DNA binding and recombination. J Mol Biol. 284:363–384. [5] Rüfer A, Neuenschwander PF, Sauer B. 2002. Analysis of Cre-loxP interaction by surface plasmon resonance: influence of spermidine on cooperativity. Anal. Biochem. 308, 90-99. [6] Hamilton D, Abremski K. 1984. Site-specific recombination by the bacteriophage P1 lox-Cre system. Cre-mediated synapsis of two lox sites. J Mol Biol. 178:481–486. [7] Hoess R, Wierzbicki A, Abremski K. 1990. Synapsis in the Cre-lox site-specific recombination system, p 203–213. In Structure & Methods. Adenine Press, New York. [8] Lee G. and Saito I. 1998. Role of nucleotide sequences of loxP spacer region in Cre-mediated recombination. Gene. 216:55±65. [9] Atasoy D, Aponte Y, Su HH, Sternson SM. 2008. A FLEX switch targets Channelrhodopsin-2 to multiple cell types for imaging and long-range circuit mapping. J Neurosci. 28(28):7025-30. [10] Thomas S. 2015. Movement Disorders: Genetics and Models: Second Edition. Chapter 3. p. 49. 云舟生物科技(广州)有限公司 020-28069042 service@vectorbuilder.cn |

更有优质直播、研选好物、福利活动等你来!




