

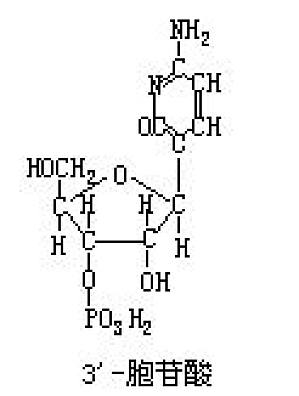
缩写CMP。为嘧啶核苷酸之一,RNA的构成成分。天然存在的有5′-胞苷酸(胞苷-5′-磷酸)。RNA碱解可生成2′-胞苷酸(胞苷-2′-磷酸)和3′-胞苷酸(胞苷-3′-磷酸)。 ...

构成核酸的嘧啶核苷之一。由胞嘧啶(碱基部分)和核糖(糖部分)组成。是通过核糖核酸水分解而成,用亚硝酸作用时,可形成尿(嘧啶核)苷。
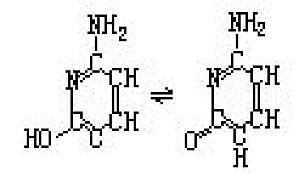
2-羟基-6-氨基嘧啶。为构成核酸的嘧啶碱基之一。系由核酸水解而成。显示特有的紫外线吸收(最大约为274纳米),可由亚硝酸的作用脱氨基变成尿嘧啶,结构如图所示,是一个互变异构体。 ...
亦称血纤维蛋白溶酶(fibrinolysin)。是存在于脊椎动物血浆中的一种蛋白酶。EC3.4.21.7.除能分解血纤维蛋白凝块使之成为可溶性外,还能使血纤维蛋白原、第Ⅴ因子、第Ⅷ因子、第Ⅶ因子,酪蛋白和明胶等蛋白质分解,并在对甲苯磺胺酰-L-精氨酸甲酯(TAMe)和赖氨酸乙酯 (LEe)其他合成底物中,水解和赖氨酸有关的多肽和酯(纤维素溶解现象)。纤维蛋白溶解活性最适的pH为7.4—7.8。 ...
在细胞组织间随着生长而产生的空隙。植物在幼嫩的时期,由于细胞互相连接,紧密地排列在一起,所以看不到胞间隙。胞间隙通常存在于已成熟的薄壁组织中,有时它们逐渐变大,同时互相连合起来,形成大的空腔或海绵状结构。虽然胞间隙按其真正的含意并不能称为组织,但它在形态、大小、内容等方面都极为多样,而且起着象细胞本身那样的作用,所以为了方便而把它作为一种组织来看待。含空气的间隙称为通气组织,含树脂、油滴等的间 ...
又称细胞间桥(cell bridge)。是指高等植物细胞间相互联系的原生质丝。根据电子显微镜的观察,它是被单位膜包围着的直径均一的(约40纳米)管状原生质,其中央有电子密度很高的物质。其外侧的膜连接在两侧细胞的细胞膜上,内部的小管也常常连接在两侧细胞的液泡系上,大小分子和病毒的核酸可通过这个结构在细胞间移动。由于胞间连丝的存在,植物体可以看作是由深深凹陷的细胞壁所支撑的巨大的多核体。在生殖细胞 ...
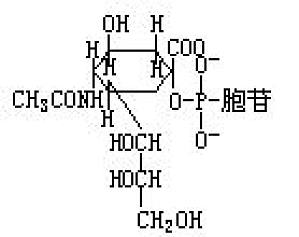
简写为CMP-N-乙酰神经氨酸。N-乙酰神经氨酸是一种多糖,特别是其作为分泌在动物细胞表面或细胞外的糖蛋白质和糖脂质末端的糖成分而广泛地分布着。其末端残基是通过总称为唾液酸转移酶(sialyltransfer-ase)的酶从这种CMP-N-乙酰神经氨酸转移而来。一般具有这种机能的糖核苷酸,是糖结合到核苷二磷酸上,但仅CMP-N-乙酰神经氨酸是例外。CMP-N-乙酰神经氨酸是通过专性的焦磷酸化酶 ...
缩写为CTP,经UTP氨化由酶催化合成,是一种在胞苷的核糖-5′-OH基上结合三分子磷酸的核苷酸。有二个高能磷酸键。是RNA生物合成的直接前体之一。参与某种多糖的合成。通过 UTP的氨基化为酶所合成。在生物合成卵磷脂、磷脂酰乙醇胺等的磷脂质时,CTP与磷酸胆碱、磷酸乙醇胺作用,酶促生成胞苷二磷酸胆碱和胞苷二磷酸乙醇胺。 ...
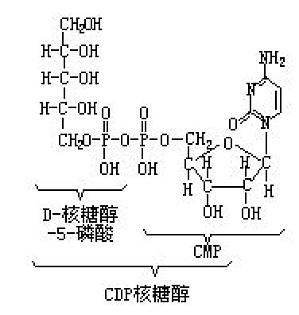
通常简写成CDP核糖醇。具有D-核糖醇-5-磷酸和 CMP以焦磷酸键连接成的结构,作为磷壁酸的多聚核糖醇磷酸键的生物合成前体,在革兰氏阳性细菌中可检测到。在由CDP核糖醇焦磷酸化酶(EC2.7.7.40)作用下通过这样的反应过程,即CTP+D核糖醇- 5-磷酸CDP核糖醇 PPi进行生物合成。以CDP焦磷酸合成多聚核糖醇磷酸的酶活性,存在于细菌的颗粒部分,但其详细的反应结构尚欠明。 ...
缩写为CDP甘油。与胞苷二磷酸核醇并列,作为磷壁酸(teichoic acid)的生物合成前体物质,分布在各种细菌中。是由甘油的1位的一级醇残基和CDP末端磷酸残基以酯键结合而成的,是通过CDP甘油焦磷酸化酶的作用以 CTP 甘油-1-磷酸CDP甘油 PPi这样的反应进行生物合成的。由此CDP甘油转移甘油磷酸而形成磷壁酸的多聚甘油磷酸骨架。 ...
CDP-OCH2CH2N (CH3)3 为动物和酵母菌等生物合成卵磷脂的中间产物。在磷酸胆碱转胞苷酰酶(EC2.7.7.15)催化下由磷酸胆碱与 CTP合成。在胆碱磷酸转移酶(cholinephoshotransferase,EC2.7.8.2)催化下,CDP胆碱与L-1,2-二甘油酯反应,产生卵磷脂。 ...
在细菌细胞表面认为有若干高分子层,而细菌细胞之所以能耐受强渗透压并能保持其独特的形状,就是因为有一种被称为细胞壁(cell wall)或者细胞囊(cell sacculus)的层包围着细胞。这个层是一个袋形的巨大分子结构体,称为胞壁质。胞壁质在几乎所有的细菌中虽有细微结构上的差异,但基本上都具有极为相似的结构。肽聚糖(peptide glycan)单位多数依靠连接其肽链部分的桥接结构排列起来, ...
3-O-α-羧乙基-D-葡萄糖胺。为D-葡萄糖胺与D-乳酸的3-O-醚。是构成细菌细胞壁的胞壁的肽聚糖的氨糖的一种。 ...
亦称实叶、芽孢叶。是与生殖有直接关系且能产生孢子的叶子的总称,为营养叶的对应词。根据形成它的生殖器是雌的还是雄的,在蕨类分为大孢子叶和小孢子叶,在种子植物则有心皮和雄蕊等区别。通常孢子叶的形态和机能已发生变态,与普通的叶很不相同,但在蕨类植物中,也有由富有叶绿体的营养叶一部分分化而成孢子囊,而孢子成熟散发后又恢复为营养叶的种类,这种叶特称为营养孢子叶。此外,在缺乏真叶的昆布类(Laminari ...
高等植物或菌类,形成营养孢子或真孢子的过程(参见孢子、分生子、游走子)。根据形成部位的不同,而分为内生孢子和外生孢子。蕨类和苔藓类,其造孢体的一部分形成孢子囊,由于其中的孢子母细胞的减数分裂,而产生单倍体的孢子。菌类的子囊孢子、担孢子,分别在子囊和担子器中由二倍体的核经过减数分裂而形成。细菌的内生孢子与上述不同,是在增殖期终了时形成持久型细胞,其形成过程曾在芽孢杆菌属(Bacillus)被详细 ...
同型不亲和性的一种类型,亦称为整体不亲和性。有关花粉亲和性的性质,是受产生花粉的孢子体基因型的影响,在柱头上由于花粉的发芽受到抑制而产生了不亲和性。这是由于孢子体中已经产生的物质通过细胞质而影响配子(花粉)的结果。例如把产生花粉的亲本的不亲和性基因的基因型记为S1S2,把特有雌蕊的基因型记为S1S3,不论花粉的基因型是S1还是S2,在S1S3基因型的柱头上花粉的发芽总是受到抑制。这样,花粉所表 ...
1 产生孢子进行无性生殖的世代称为孢子体世代(sporophytic generation),这个世代的生物体称为孢子体。在孢子形成中,一般伴有减数分裂。与此相反,形成配子进行有性生殖的,则分别称为配子世代和配子体。孢子体(核相2n)和配子体(核相n),在生活史上交替出现的现象,称为世代交替。孢子体和配子体的形态有完全相同的,也有显著不同的。苔藓植物和维管植物均属于后者。此外,F.Mocwus ...
1 孢子虫类的合子(受精卵),在卵囊内分裂,形成许多孢子母细胞,孢子母细胞再分裂后才形成孢子,这时,把孢子母细胞在卵囊中或卵囊外形成的膜,或连同内含物一起称为孢子囊。孢子母细胞往往不经分裂,仅在外层产生被膜直接形成孢子。 2 胞蚴见于吸虫纲的复殖类发育中,是继毛蚴之后,相当于单性虫(parthenitae)的第一代。胞蚴往往见于中间宿主腹足类被毛蚴侵入部位附近,胞蚴体呈囊状,无消化器 ...
内部生有孢子的囊状生殖器官。亦称芽孢囊。内部的孢子称孢囊孢子。不同种的植物,其形状多种多样,一般当成熟时即裂开而散出孢子,也有的是由于囊的破裂或溶化而散出孢子。在同种植物中形成的孢子囊生成同样的孢子时称为同形孢子囊,如产生大小两种孢子时则称为异形孢子囊,后者分别为大孢子囊和小孢子囊,如水生蕨类和水韭。藓类的孢子囊特称为孢蒴。在种子植物中,胚珠相当于大孢子囊,花药相当于小孢子囊。苔类的也有一种孢子囊 ...
1 棘毛。 2 从子囊菌类的子囊壳或半知菌类的分生孢子器的开口处,被连续压出而形成卷须状或带状的孢子块。 ...
关于丁香通
公司信息
个人用户
企业机构


