

- 询价
- Xybio
- P4216
- 上海
- 2025年11月05日

企业认证
相关产品推荐更多 >





万千商家帮你免费找货
0 人在求购买到急需产品
- 详细信息
- 文献和实验
- 技术资料
- 库存:
100
- 英文名:
NM_004985.4;C-K-RAS;c-Ki-ras2;CFC2;K
- 保质期:
5年
- 供应商:
上海烜雅生物科技有限公司
- 保存条件:
-20℃
名称:pCMV-SPORT6-KRAS(1点突变)人源基因质粒
别称: NM_004985.4;C-K-RAS;c-Ki-ras2;CFC2;K
复制子: pUC
原核抗性: Amp
克隆菌株: DH5a
培养条件: 37度
质粒属性
载体宿主:大肠杆菌
载体用途:PCR模板
基因种属:人
基因类型:cDNA
原核抗性:Amp
真核抗性:
荧光蛋白:
质粒简介
pCMV-SPORT6-KRAS(1点突变)人源基因质粒。
质粒图谱:祥询
质粒序列:祥询
质粒菌株产品操作说明书
一、扩增流程
收到产品后,请先根据产品管壁标签来判断产品形式,并在扩增前准确查找该质粒菌株的抗性、感受态和培养温度。
1、质粒干粉(常温运输,存于-20度,90天保质期,请务必转化挑单克隆培养,不要直接使用和测序)
①收到质粒干粉后请先5000rpm离心1min,再加入20μl ddH2O去离子水溶解质粒;
②取1支100μl 感受态于冰上解冻10min,加入2μl质粒,再冰浴30min后,42℃热激60s,不要搅动,再冰浴2min;(从第二步开始均要在超净工作台中无菌操作)
③加入900μl无抗的LB液体培养基,180rpm震荡37℃培养45min;
④6000rpm离心5min,仅留100μl上清液重悬细菌沉淀,并涂布至目标质粒抗性的LB平板上;(可使用本平台的平板涂布专用玻璃珠进行涂布,可以比传统涂布方法获得更多转化子)
⑤将平板正向培养1h,再倒置37℃培养14h。如果要求是30度则培养20h;
(菌落过多则将质粒稀释后再转化。没有菌落则加入10μl质粒转化。另不要直接转表达感受态,要先转克隆感受态,重提质粒后再导入表达感受态)
⑥挑取单菌落至LB液体培养基中,加入对应抗生素,220rpm震荡培养14h,根据实验需要和质粒提取试剂盒说明书提取质粒。
2、甘油菌种(冰袋运输,存于-80℃,保质期90天,请务必划线挑单克隆培养)
四区划线后挑单菌落培养,酵母菌需要先液体复苏再四区划线,再挑单菌落液体培养。
3、穿刺菌种(冰袋运输,存于4℃,保质期7天)
穿刺接种,液体培养后四区划线,再挑单菌落液体培养。
4、菌落平板(冰袋运输,存于4℃,保质期7天)
直接挑取单菌落至液体培养基中。
5、液体质粒(冰袋运输,存于-20℃,保质期90天)
单独提取的液体质粒收到后可直接使用。
6、滤纸质粒(常温运输,存于-20度,90天保质期,请务必转化挑单克隆培养,不要直接使用和测序)
收到货后将滤纸画圈部分剪下放入EP管中,加100ul无菌水将滤纸浸湿并浸泡5min,吸取5ul质粒转化,离心全涂。
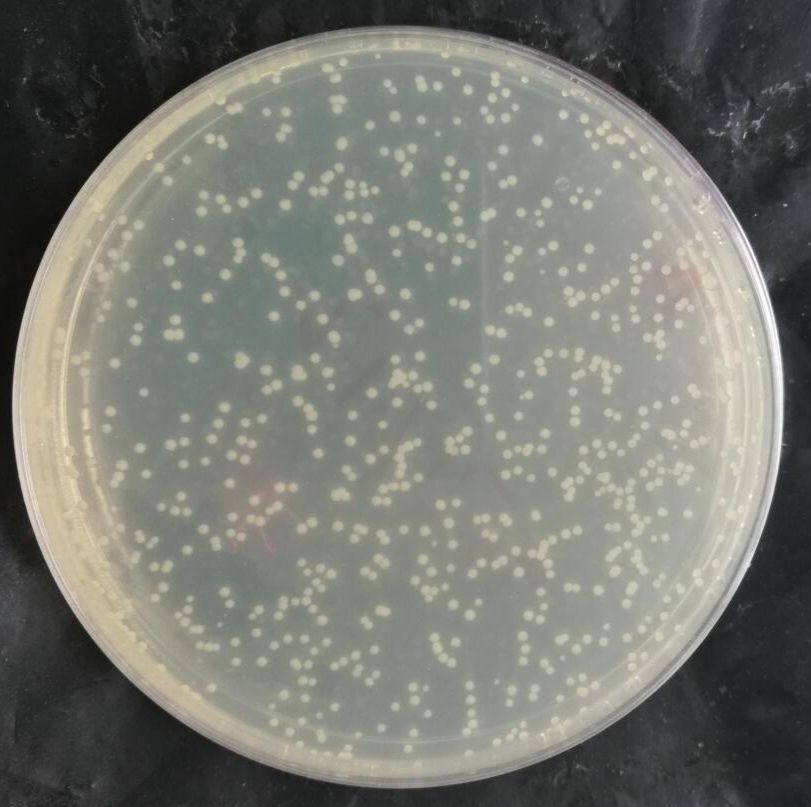
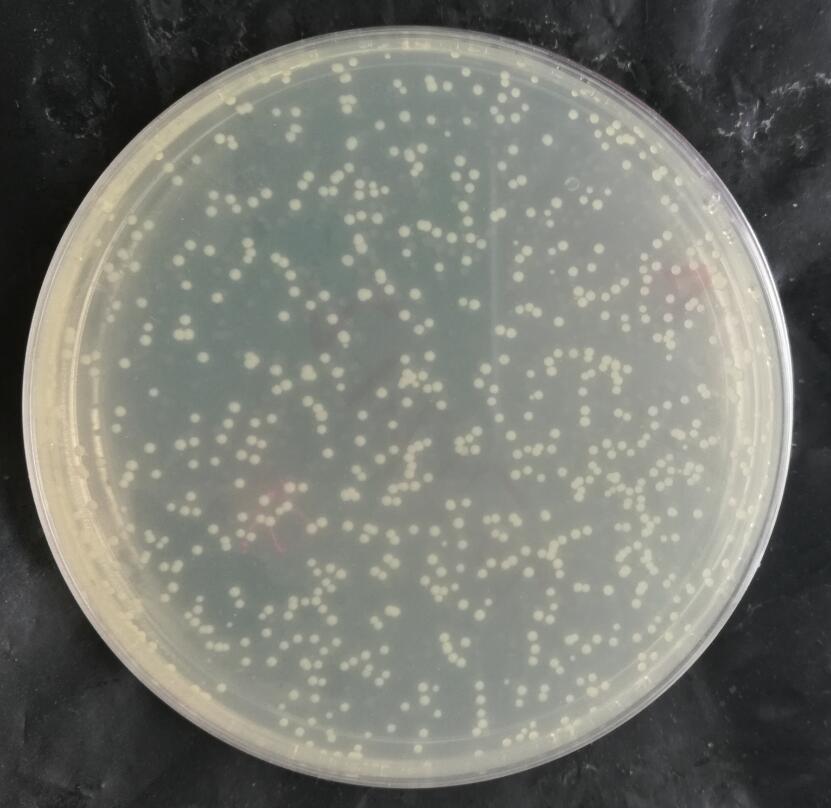
二、转化图片
相关产品
| P4072/pCMV-SPORT6-CENPH(1同义突变)人源基因质粒 |
| P4073/pCMV-SPORT6-CHURC1(10-420bp)人源基因质粒 |
| P4074/pCMV-SPORT6-TRIR人源基因质粒 |
| P4075/pCMV-SPORT6-NLRP1人源基因质粒 |
| P4076/pCMV-SPORT6-RFK人源基因质粒 |
| P4077/pCMV-SPORT6-CXCL1人源基因质粒 |
| P4078/pCMV-SPORT6-RPL35A人源基因质粒 |
| P4079/pCMV-SPORT6-CRADD人源基因质粒 |
| P4080/pCMV-SPORT6-CCDC127人源基因质粒 |
| P4081/pCMV-SPORT6-CIDEC(5-756bp1同义突变)人源基因质粒 |
| P4082/pCMV-SPORT6-SSSCA1人源基因质粒 |
| P4083/pCMV-SPORT6-AGR2(1同义突变)人源基因质粒 |
风险提示:丁香通仅作为第三方平台,为商家信息发布提供平台空间。用户咨询产品时请注意保护个人信息及财产安全,合理判断,谨慎选购商品,商家和用户对交易行为负责。对于医疗器械类产品,请先查证核实企业经营资质和医疗器械产品注册证情况。
 文献和实验
文献和实验),绿色荧光蛋白(GFP),荧光素酶(Lux或Luc)以及b-半乳糖苷酶(b-gal)。可以使用简单的非同位素方法检测b-gal的表达以测定转染效率和活性。pCMV SPORT- bgal质粒包含CMV启动子调控下的LacZ基因,转染入真核细胞内后可以直接表达bgal。结合简单的检测步骤,可以做为监测转染条件的一种方便灵敏的方法。 09稳定转染细胞系的筛选 用载体中所含的选择标志进行筛选是建立稳定转染细胞系最常用的方法。抗生素抗性基因可以与目的基因在同一个质粒上,也可以在不同的质粒上。如果两个不同
-crRNA 和加长 pre-crRNA 融合构建了 crRNA-Cpf1 系统,在水稻中的编辑效率最高可达 41.2%;HU 等也将 LpCpf1 和 crRNA 整合开发了 CRISPR-Cpf1 系统,并利用该系统成功实现水稻内源基因的定点编辑;KIM 等开发了 Cpf1–RNP(ribonucleoprotein) 系统,并成功利用该系统对大豆和烟草的内源基因进行定点突变。此外,BEGEMANN 等将 Cpf1、crRNA 和供体片段构成融合表达载体,对水稻叶绿素 a 加氧酶基因 OsCAO
的上下游两端的序列连接起来,从而实现了目的基因的敲除。再如,可为细胞提供一个修复的模板质粒,这样细胞就可按照提供的模板在修复过程中引入片段插入或定点突变。也可对受精卵细胞进行基因编辑,并将其导入代孕母体中,可实现基因编辑动物模型的构建。


