- 移动端


江苏赛昂斯生物科技有限公司
1 年
手机商铺
- NaN
- 0.2999999999999998
- 0.2999999999999998
- 2.3
- 2.3
推荐产品




技术资料/正文
Morris水迷宫 | 逆转由慢性皮质酮诱导的焦虑样抑郁行为:西红花素I的干预作用及血浆代谢物的表面增强拉曼光谱监测
561 人阅读发布时间:2025-10-09 16:35

论文上线截图
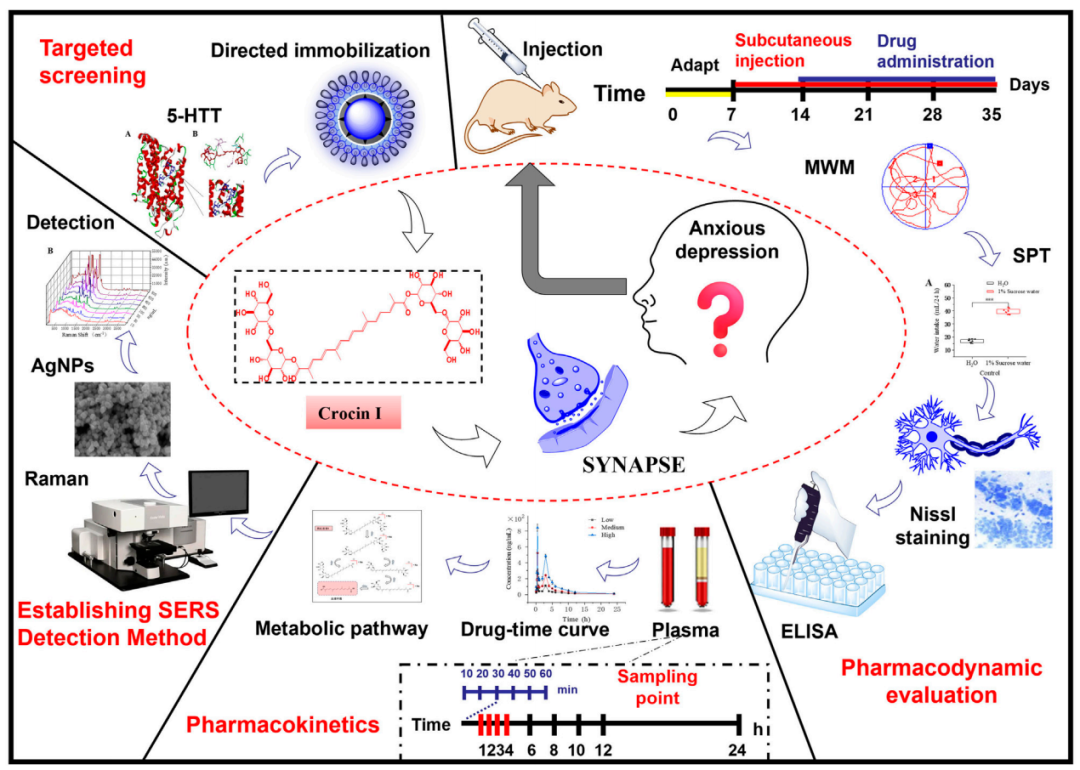
研究首次发现栀子中的主要成分藏红花苷Ⅰ(crocin I)能够逆转慢性皮质酮诱导的大鼠焦虑样抑郁行为及海马形态学改变,显著修复海马齿状回、CA1和CA3区的神经元损伤,并通过调节脑内单胺类神经递质(5-HTT、DA、NA)和GABA水平发挥抗焦虑抑郁作用。药代动力学分析显示,crocin I自身难以通过肠道屏障进入血液,其代谢产物藏红花酸(crocetin)可被吸收并发挥作用。创新性地,研究团队利用表面增强拉曼光谱(SERS)技术,建立了高灵敏度、快速检测血浆中微量藏红花酸的方法,提升了对其体内代谢的监测能力。研究不仅揭示了crocin I治疗焦虑抑郁的潜力和机制,也为其临床应用及相关天然药物的开发提供了理论基础和技术支持。
1. 藏花素I能以剂量依赖的方式显著改善焦虑样抑郁大鼠的体重减轻和快感缺失行为
实验初期大鼠因环境适应出现体重下降,随后数周体重增长,但在第五周因水迷宫实验压力导致全体重下降,其中低剂量组和模型组体重增幅小于其他干预组(图1A-C)。模型组大鼠的糖水偏好指数(43.3%)显著低于对照组及所有干预组,而各给药组均表现出不同程度的糖偏好改善,且对藏花素I的反应呈剂量依赖性(图1D-K)。综上所述,藏花素I能有效改善焦虑样抑郁大鼠的体重减轻和快感缺失行为。
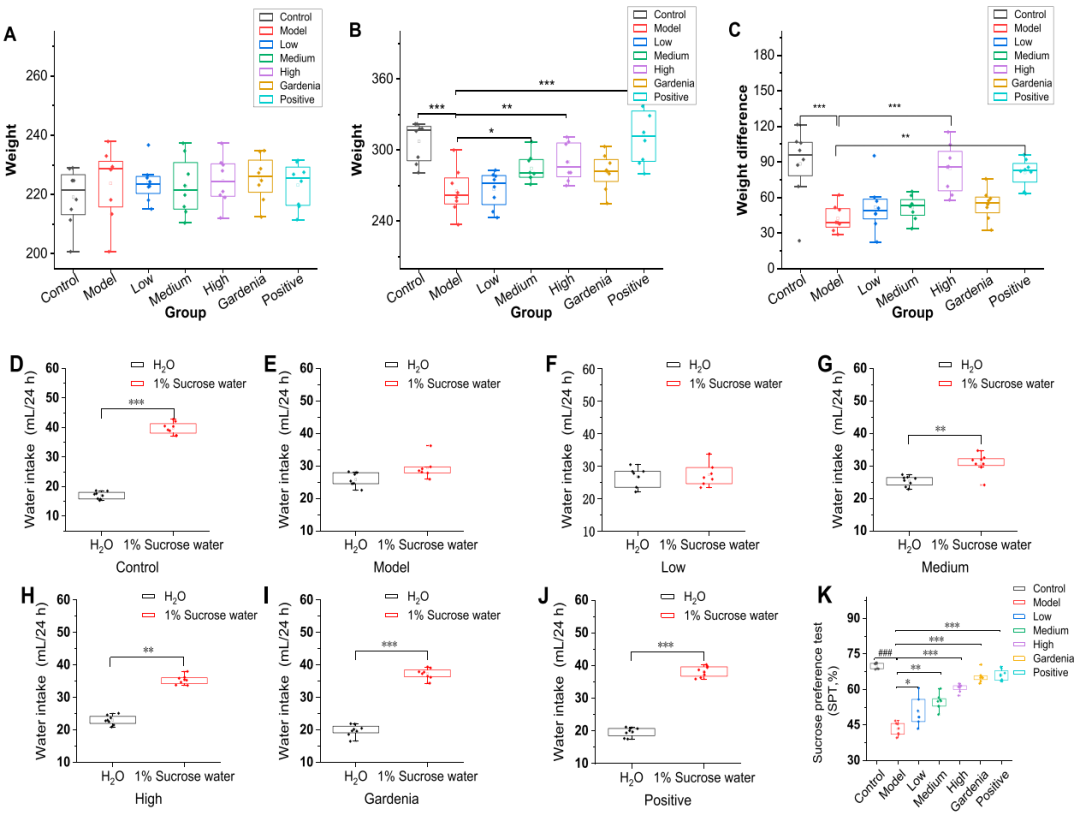
图1.各组大鼠体重差异及糖水偏好率分析
2. 西红花苷I能够以剂量依赖的方式显著改善焦虑样抑郁大鼠的空间学习与记忆功能
水迷宫测试结果显示,模型组大鼠表现出明显的空间记忆与认知能力受损,具体表现为逃避潜伏期延长(图2I)、穿越目标象限次数减少、运动距离缩短,并更多采用边缘式搜索策略(图2A-B)。而经西红花苷I干预(图2C-G)后,各剂量组大鼠均呈现剂量依赖性的改善作用,表现为逃避潜伏期显著缩短(图2I)、目标象限距离占比升高(图2H),搜索策略由边缘式逐步转变为趋向式(图2A-C)。此外,西红花苷I组与阳性药物帕罗西汀组均显示出与模型组和对照组相比具有显著差异的行为恢复。因此,西红花苷I以剂量依赖的方式显著改善了由皮质酮诱导的焦虑样抑郁大鼠的空间学习与记忆能力,具体表现为缩短逃避潜伏期、增加目标象限穿越次数及促进更高效的搜索策略。
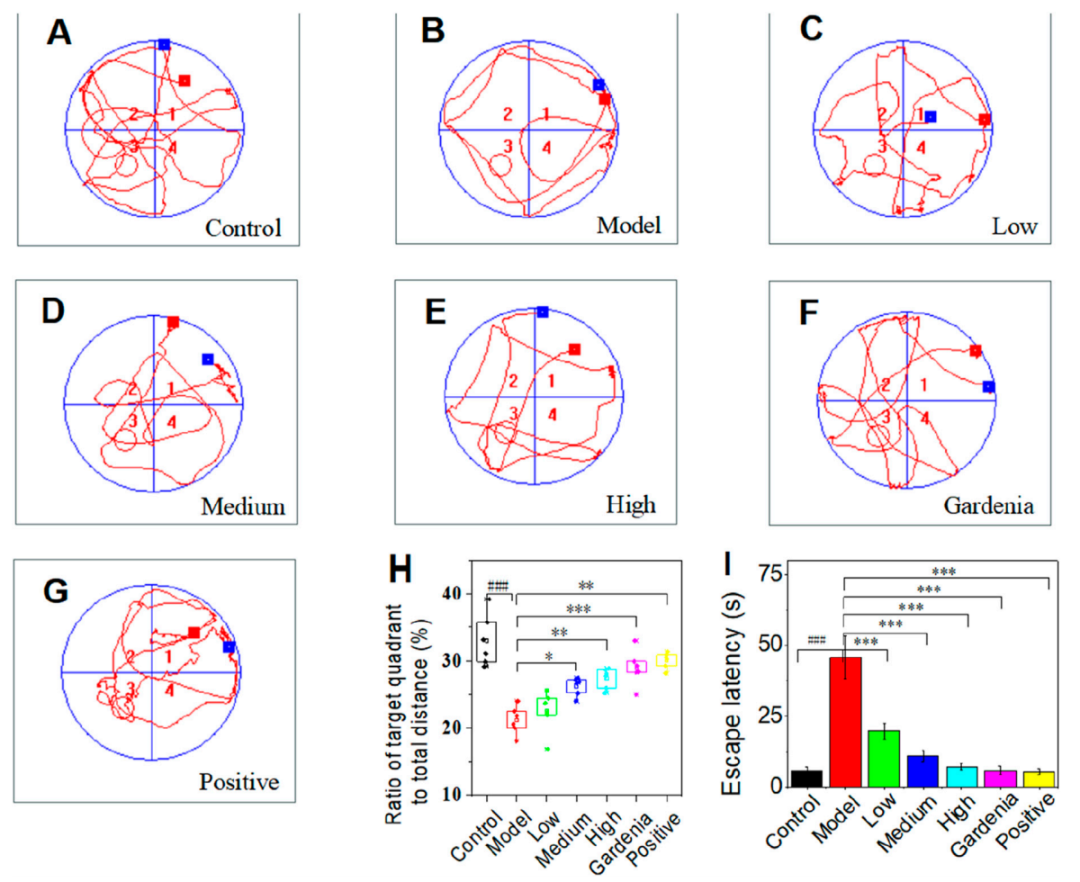
图2.各组大鼠水迷宫空间探索轨迹图
3. 西红花素I能剂量依赖性地改善海马神经元结构与形态损伤,其高剂量组与栀子提取物组对齿状回和CA1区的保护作用尤为显著
模型组海马齿状回边缘钝化、神经元边界模糊且尼氏体减少,而西红花素I中高剂量组和栀子提取物组能显著改善齿状回形态锐度、恢复神经元结构完整性并增加尼氏体数量,其中高剂量西红花素I与栀子提取物效果尤为显著,阳性药物组虽能改善神经元损伤但对形态学变化干预有限(图3A-G)。总结而言,西红花素I通过剂量依赖性方式有效逆转皮质酮诱导的海马结构与神经元损伤,其疗效优于常规阳性药物。
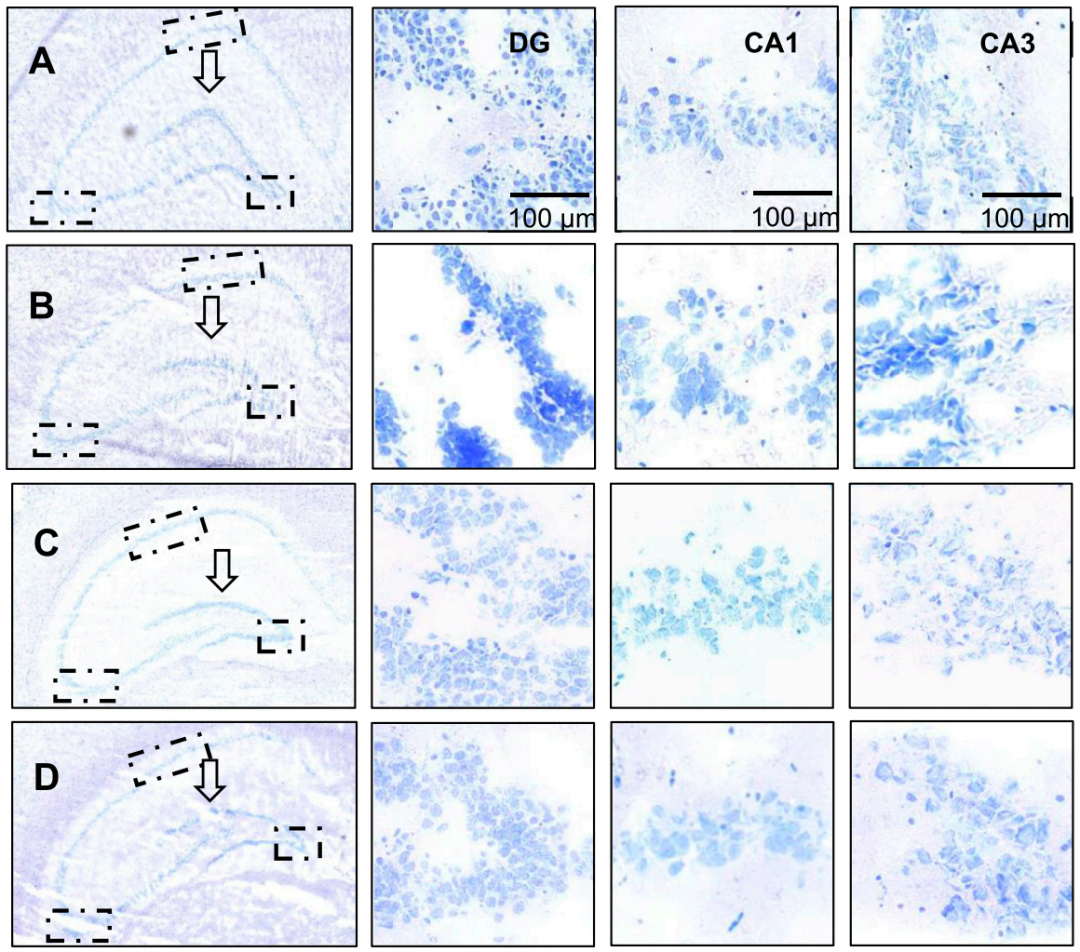
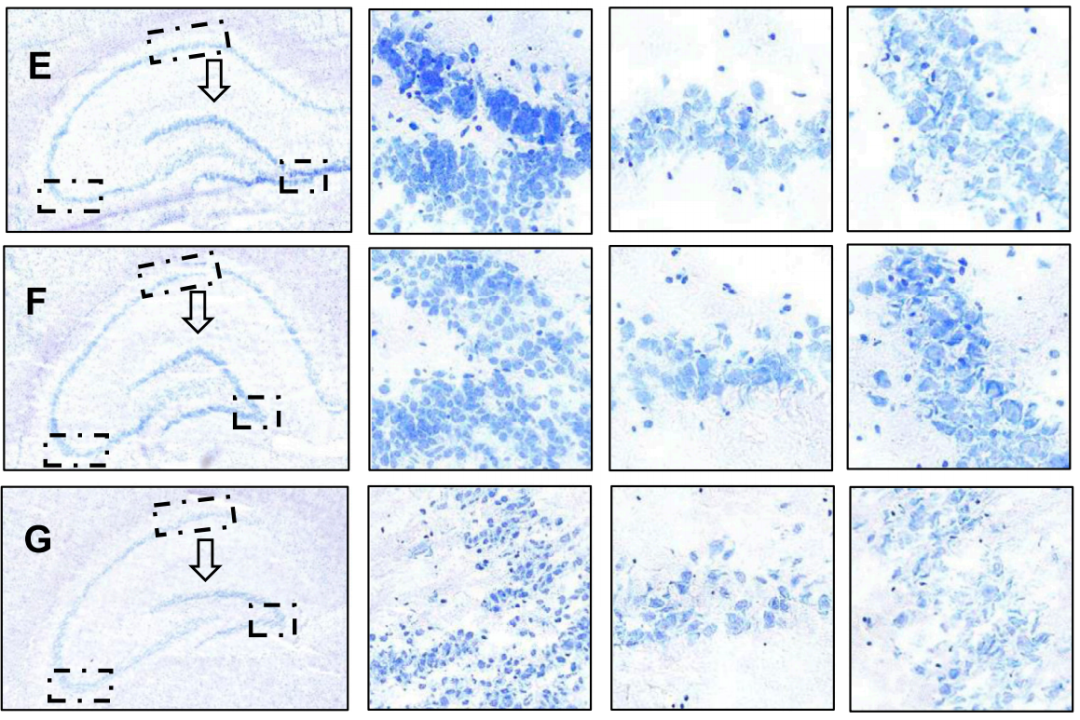
图3.大鼠海马神经元尼氏染色分析
4. 西红花素I、栀子提取物及阳性药物均能显著改善模型大鼠皮质神经元损伤,增加尼氏体数量,显现明确的神经保护作用
与结构规整、尼氏体丰富的对照组(图4A)相比,模型组大鼠前额叶皮层神经元出现核固缩、尼氏体溶解等显著损伤(图4B);而经高剂量西红花素I、栀子提取物及阳性药物治疗后,神经元尼氏体着色加深、密度增大,细胞损伤得到显著修复(图4C-G),且通过定量分析证实各治疗组均能有效增加模型大鼠皮质神经元中尼氏体的数量(图4H)。总而言之,该实验结果表明西红花素I、栀子提取物及阳性药物均对皮质神经元损伤具有明确的保护作用。
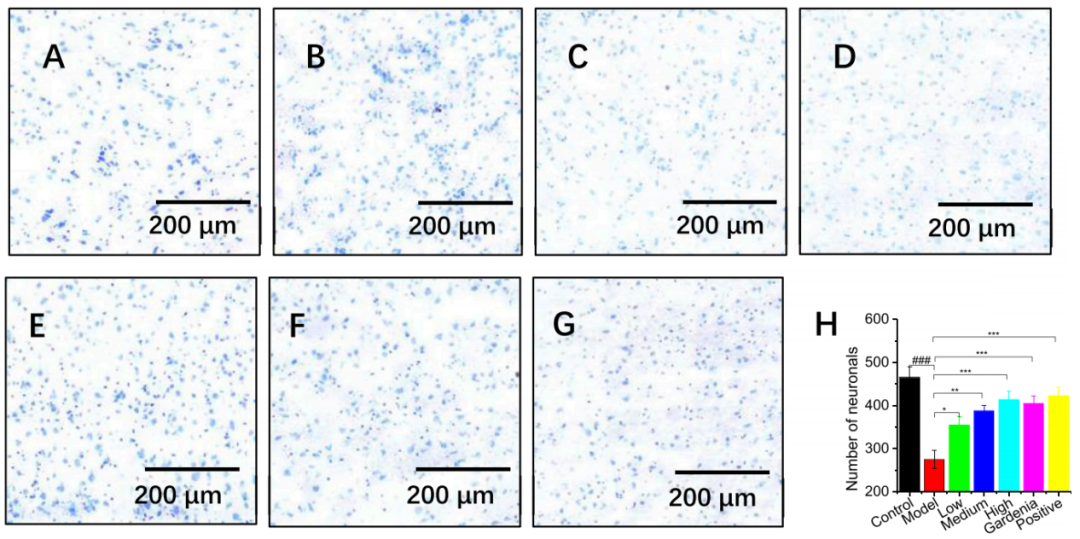
图4.不同组别大鼠对前额叶皮层的影响
5. 藏红花素I与栀子提取物通过显著调节5-HTT、GABA、DA及NA水平,有效改善了抑郁模型大鼠的单胺类神经递质紊乱
与模型组相比,藏红花素I各剂量组、栀子提取物组及阳性药物组均能显著降低焦虑抑郁大鼠脑中过度表达的5-HTT含量,并呈现剂量依赖性(图5A);同时,中、高剂量藏红花素I、栀子提取物及阳性药物能显著升高异常的GABA水平(图5B);而与对照组相比,模型组组大鼠脑内多巴胺(DA)和去甲肾上腺素(NA)含量显著升高,藏红花素I高剂量组、栀子提取物组及阳性药物组的大脑DA含量与对照组相比无显著差异 (图5C-D)。综上,这些结果证明藏红花素I和栀子提取物能有效调节焦虑抑郁模型大鼠脑内的单胺类神经递质紊乱。
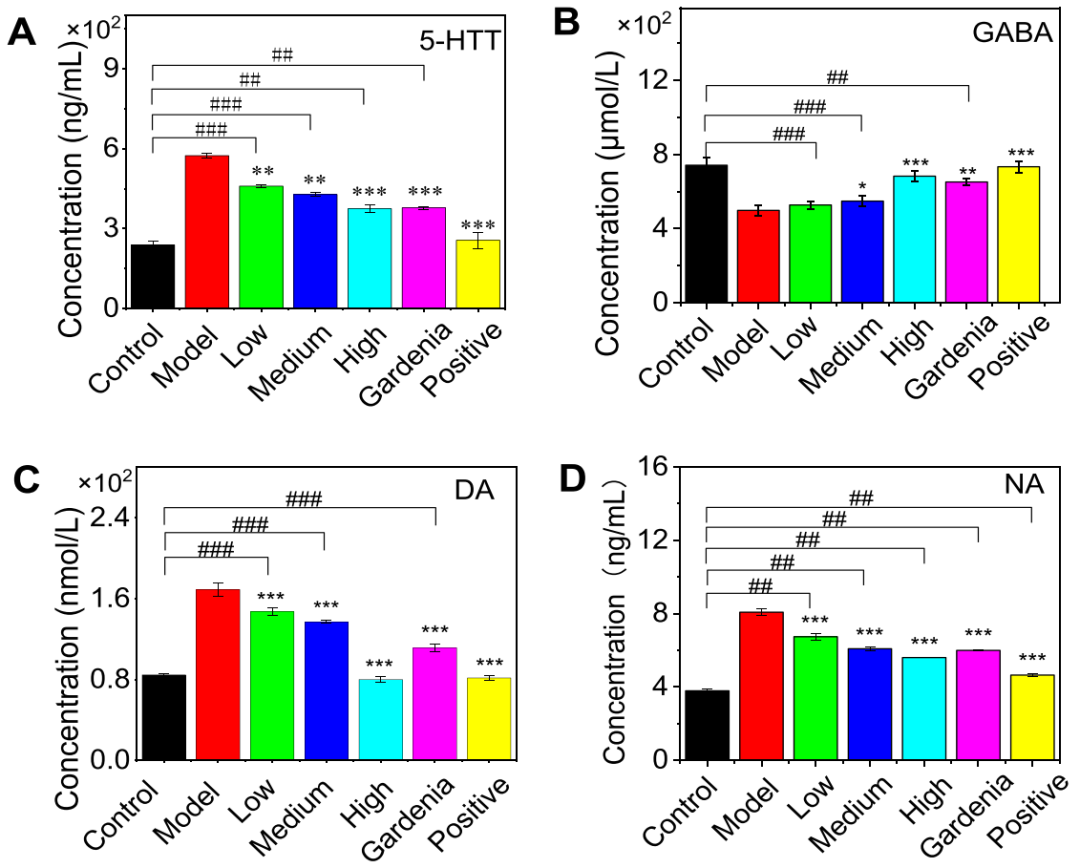
图5.西红花苷Ⅰ和栀子提取物对焦虑抑郁模型大鼠脑内5-HTT、GABA、DA、NA表达的影响
6. 西红花苷I经代谢转化为西红花酸,后者能有效穿透肠道屏障进入血液,这是其实现生物利用的关键途径
如图6所示,实验结果表明西红花苷I在体内代谢后主要生成西红花酸,且该代谢产物能有效穿透肠道屏障进入血液系统(图6)。结果表明,西红花苷I是通过转化为西红花酸实现其生物利用度的关键途径。
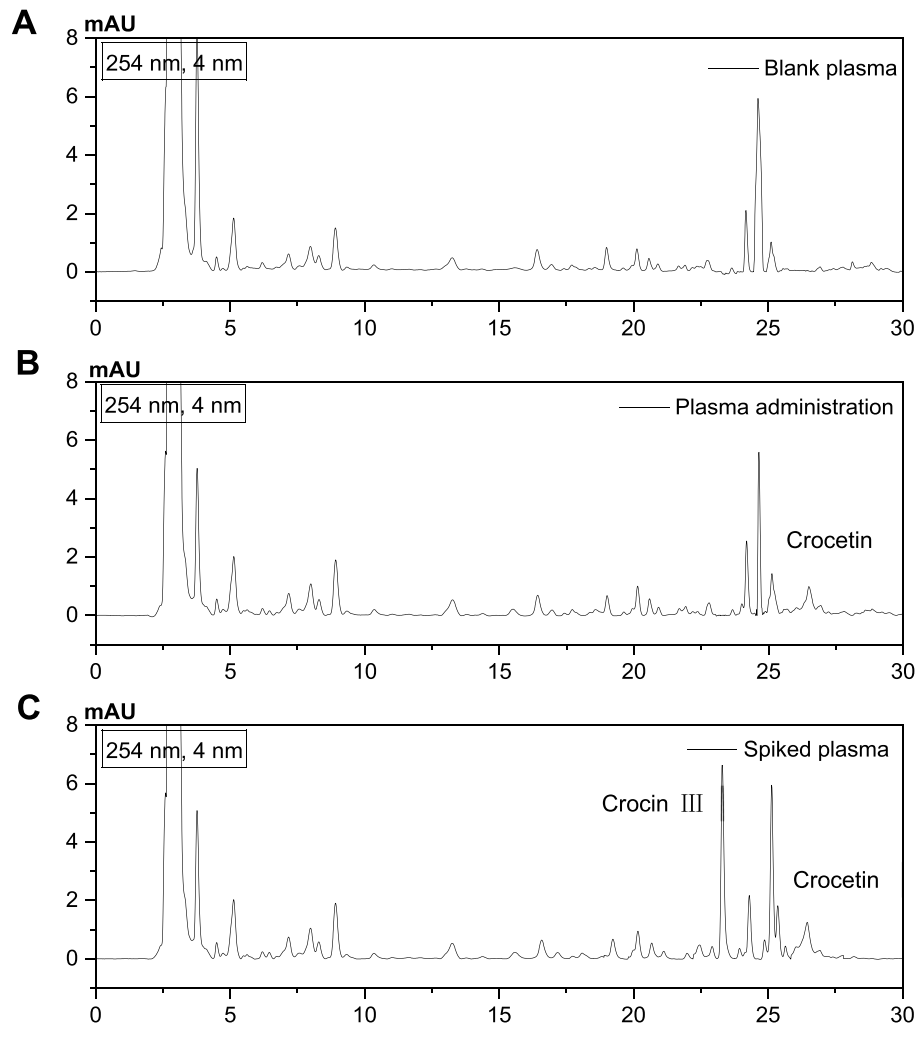
图6.通过HPLC分析代谢物
7. 西红花苷Ⅰ的代谢终点是生成分子结构更为简单的西红花酸
西红花苷Ⅰ在体内通过逐步裂解糖苷键,依次生成多种代谢产物,并最终代谢为仅含四个异戊烯基结构的最终产物西红花酸 (图7)。该结果清晰地揭示了西红花苷Ⅰ在生物体内的完整代谢路径。
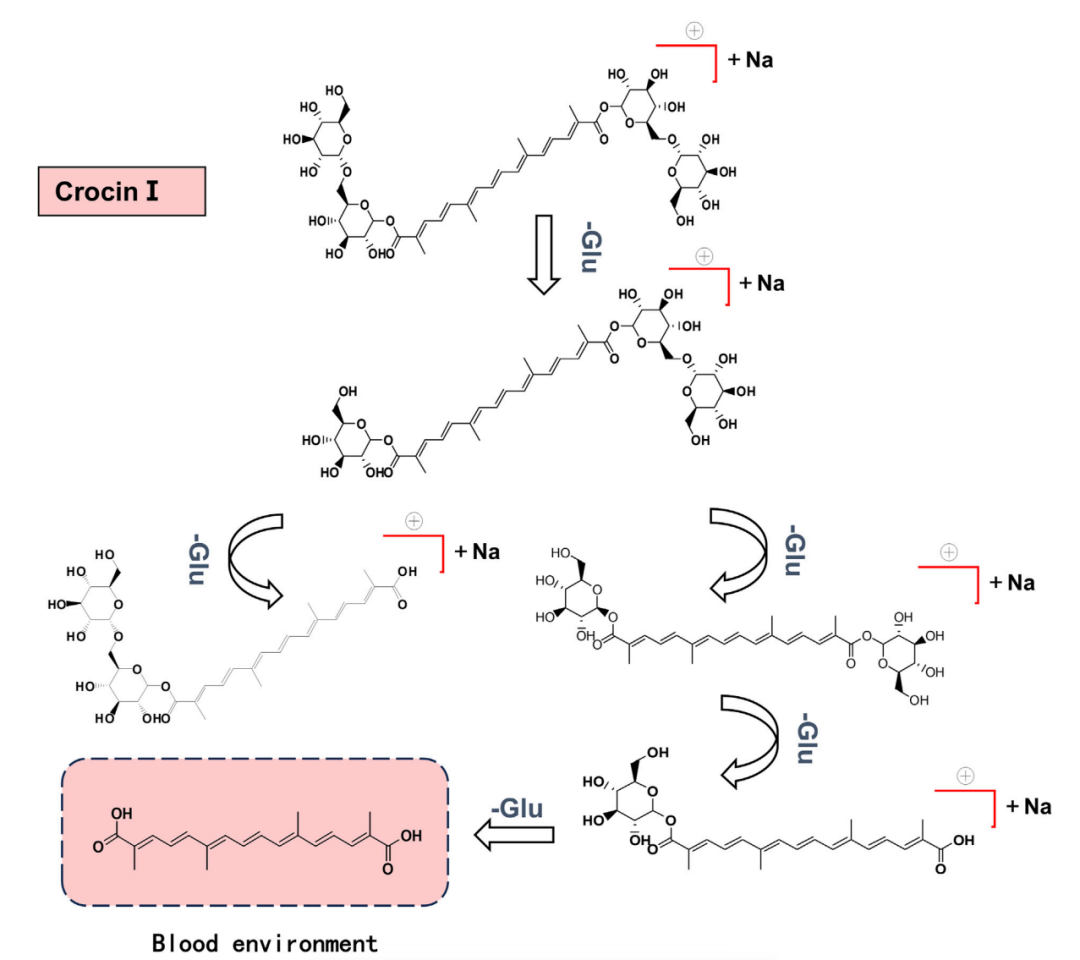
图7.大鼠西红花苷Ⅰ裂解途径
8. rocin I在大鼠体内呈现多峰吸收现象,表明其胃肠道吸收过程存在复杂性
如图8所示,代谢产物Crocin I在大鼠体内不同时间点出现多个血药浓度峰值,提示其吸收过程可能受胃内吸收位点差异、肠肝循环或胃液pH与胃排空速率不匹配等因素影响。总结来说,该药代动力学曲线呈现出多峰特征,表明其体内吸收过程较为复杂。
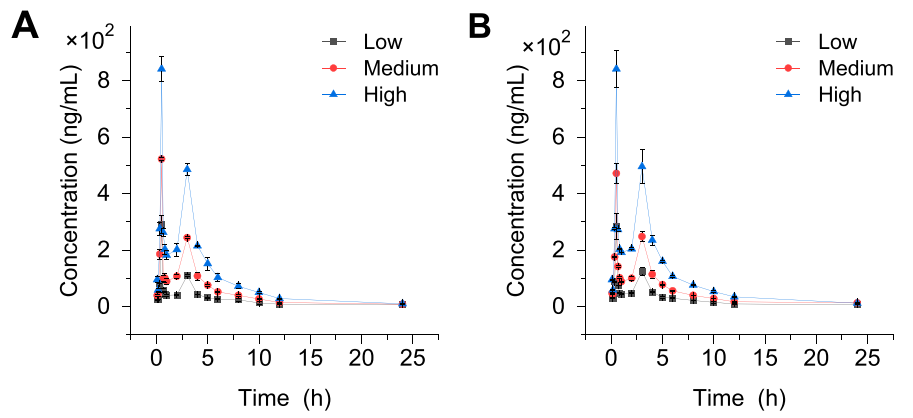
图8.造模前后的药时曲线
9. 合成的银纳米粒子能显著增强血浆中藏红花酸的拉曼信号强度,为其高灵敏度检测提供有效方法
通过扫描电镜观察(图9A, B)证实成功合成了平均粒径为81.05 nm的银纳米粒子,其拉曼光谱见图9C;实验进一步利用该基底材料检测血浆中的藏红花酸,结果显示未经纳米银增强时,藏红花酸的特征拉曼峰信号极弱甚至不可见(图9D, E),而经增强后这些特征峰的信号强度和峰形均得到显著改善(图9F)。综上所述,该研究表明所合成的银纳米粒子能有效增强藏红花酸的拉曼信号,可用于其高灵敏度检测。
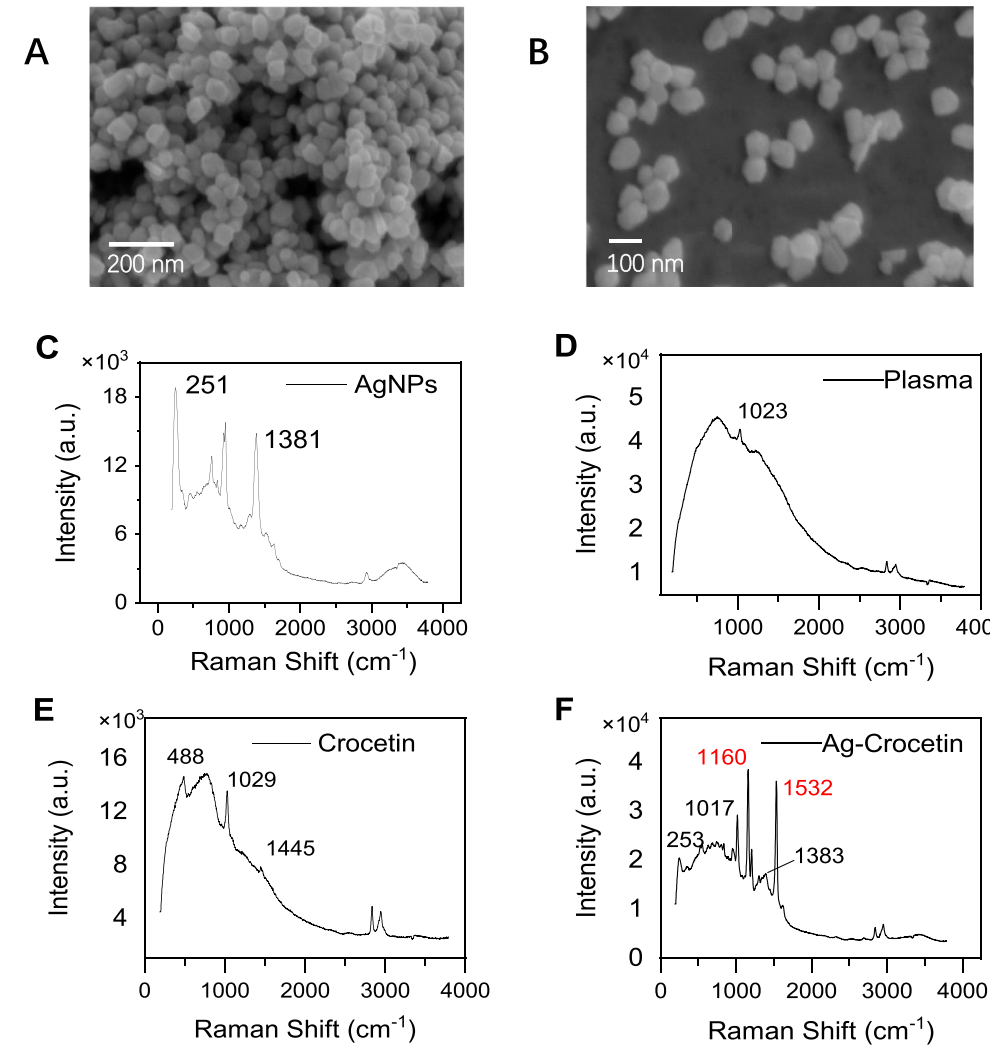
图9.使用扫描电子显微镜(SEM)和拉曼光谱法。
10. SERS方法具有稳定的检测性能、高灵敏度(10 ng/mL)和良好的定量能力,适用于西红花酸的快速分析
该SERS检测方法在一周内保持光谱稳定性(图10A),在10-800 ng/mL范围内呈现良好线性关系(图10C),最低检测限达10 ng/mL(图10B),血浆样本定量结果与液相方法吻合良好(图10D)。该SERS方法具有优异的稳定性、灵敏度和定量检测能力,完全满足西红花酸快速分析要求。
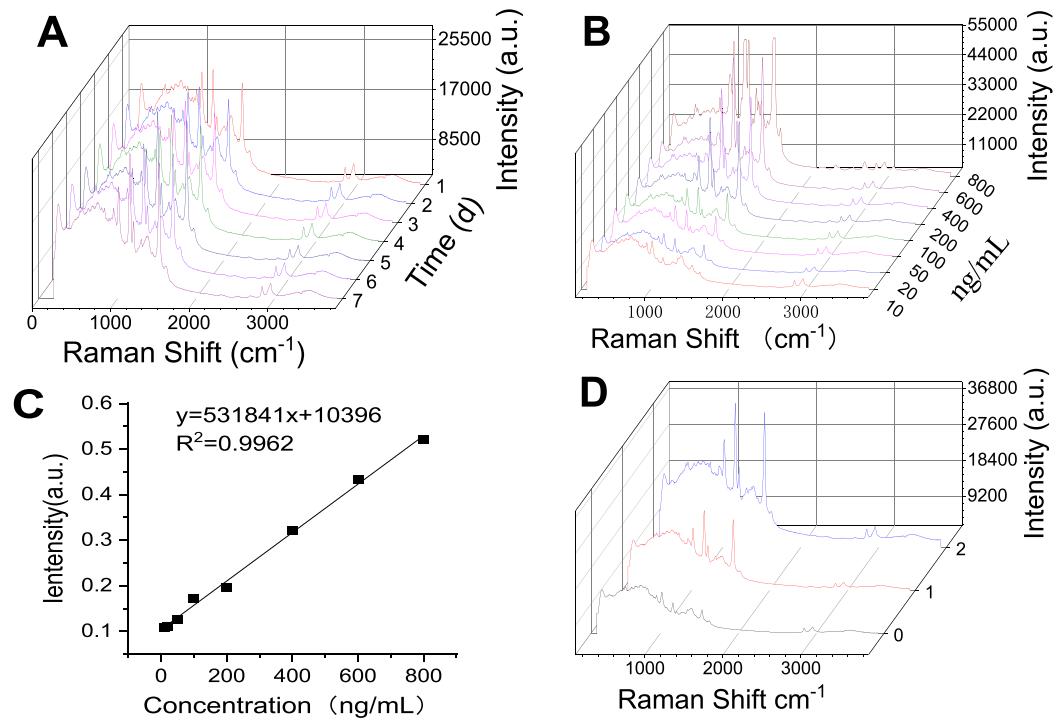
图10.藏红花酸的表面增强拉曼检测方法学研究
结论
研究首次揭示了藏红花成分crocin I可显著逆转慢性皮质酮诱导的大鼠焦虑抑郁行为及海马神经元损伤,并通过行为学与组织学实验证实其改善认知情绪、修复海马DG/CA1/CA3区形态的作用。机制上,crocin I调控单胺类神经递质(如5-HTT、DA、NA和GABA)表达,抑制5-羟色胺转运体过度活性,恢复神经传导功能。药代动力学研究表明crocin I主要以其代谢产物crocetin发挥系统作用,研究团队进一步开发了基于银纳米颗粒的表面增强拉曼散射(SERS)技术,实现了血浆中crocetin的高灵敏检测(检测限10 ng/mL,增强因子4.49×10^11),为药物体内监测与临床应用提供了重要技术支撑。该研究不仅证实crocin I的神经保护与抗焦虑抑郁潜力,也推动了天然产物在精神疾病治疗中的开发与应用。
SA201 Morris水迷宫
Morris水迷宫(Morris water maze, MWM)实验是一种强迫实验动物(大鼠、小鼠)游泳,学习寻找隐藏在水中平台的一种实验,主要用于测试实验动物对空间位置感和方向感(空间定位)的学习记忆能力。被广泛应用于学习记忆、老年痴呆、海马/外海马研究、智力与衰老、新药开发/筛选/评价、药理学、毒理学、预防医学、神经生物学、动物心理学及行为生物学等多个学科的科学研究和计算机辅助教学等领域,在世界上已经得到广泛地认可,是医学院校开展行为学研究尤其是学习与记忆研究的首选经典实验。

SA226 糖水偏好实验系统
糖水偏好实验系统有实时统计、自动化、高准确性等优点,极大地提高了药物研发和基础生命科学研究的效率,并从根本上减少手工操作带来数据偏差及误差。
在动物无拘束状态下,进行多通道测量,记录软件能实时统计大小鼠的饮水次数,饮水时间,饮水量等多个指标,饮食槽设计防止粪便的混入及饵料的抛洒,粪尿完全分离。可做到短期、长期多通道监测。
