- 移动端


江苏赛昂斯生物科技有限公司
1 年
手机商铺
- NaN
- 0.2999999999999998
- 0.2999999999999998
- 2.3
- 2.3
推荐产品




技术资料/正文
睡眠剥夺仪 | 在急性睡眠剥夺及短期睡眠恢复期间持续的小胶质细胞激活和树突棘的加速消除
250 人阅读发布时间:2025-01-15 12:56

论文上线截图
睡眠是一种由多种神经解剖学和神经化学系统严格调节的生理状态。以往研究表明,补觉对于抵消睡眠负面影响的效果有限,然而其背后的机制尚不清楚。在这项研究中,作者使用了多种转基因荧光小鼠模型以及活体双光子成像和脑切片免疫荧光染色等技术,记录了急性睡眠剥夺(ASD)及随后的短期恢复睡眠(RS)期间小胶质细胞的形态和功能变化,及其对神经元突触结构可塑性的效应,并结合全脑组织代谢组分析,揭示了睡眠不足对神经免疫调节和神经可塑性的不利影响,并探讨了不规律睡眠导致神经系统疾病的潜在机制。
1. 在ASD及RS期间,小胶质细胞持续激活
在健康大脑中,小胶质细胞通常处于生理状态,其不断地监测大脑实质的周围微环境,寻找细胞外信号,并与其他细胞包括神经元和星形胶质细胞进行互动。然而,这种稳态可能会受到外部刺激和病理生理条件的干扰,表现出小胶质细胞“激活”的不同状态。为了记录ASD期间及随后的RS期间不同大脑区域中小胶质细胞的动态形态变化,作者观察了ASD和RS组(图1)小鼠的脑切片,通过双光子成像技术对大脑皮层、海马CA1、齿状回多形层(PoDG)以及纹状体区域的小胶质细胞进行了成像(图2A),发现在ASD 48h和RS 24h组中,小胶质细胞表面积、过程长度和小胶质细胞过程端点数量显著且持续减少(图2B-D)。以上结果表明,在ASD及随后的短期RS期间,小胶质细胞的激活持续处于低水平。
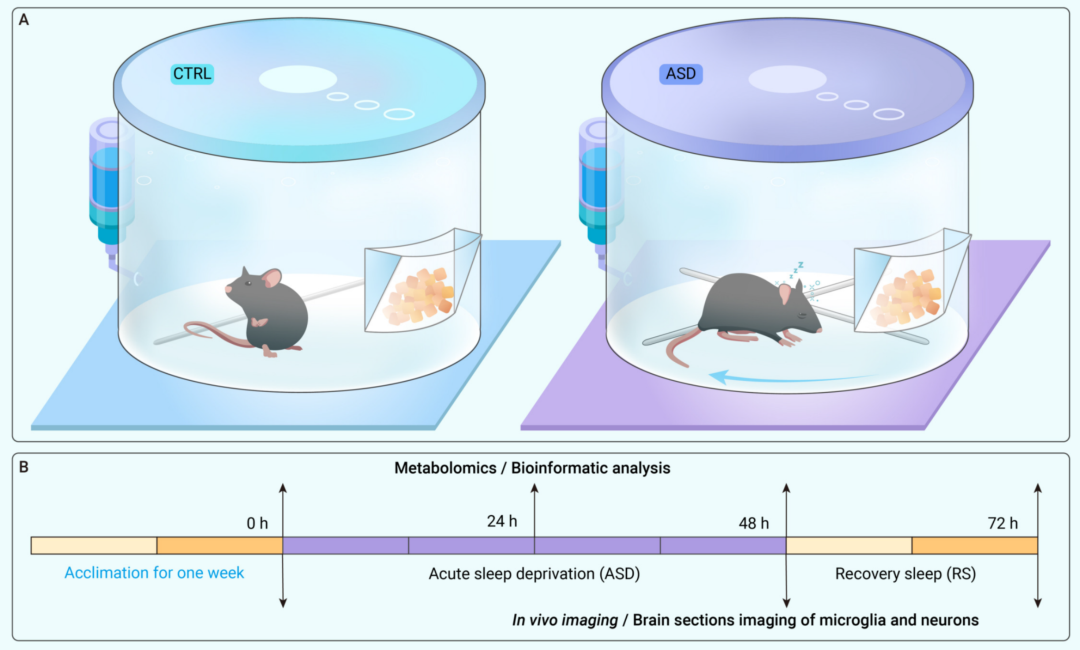
图1. 急性睡眠剥夺(ASD)模型及实验设计
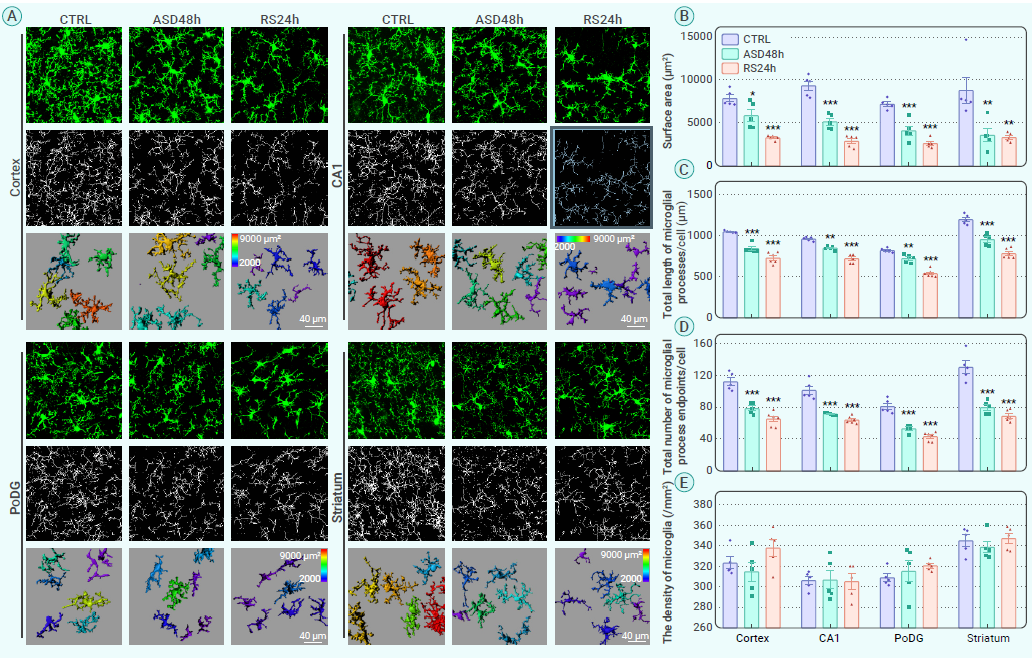
图2. 小胶质细胞在ASD和随后的RS期间显示持续激活
2. 在ASD和RS期间,小胶质细胞突起的活动持续降低
作为中枢神经系统中的监视免疫细胞,小胶质细胞通过其突起活动的动态过程响应微环境的变化。尽管小胶质细胞突起活动重要,但这些过程在不同生理条件下的变化仍不太清楚。作者通过活体双光子成像技术,研究了ASD和随后RS期间躯体感觉皮层中小胶质细胞突起的活动(图3A)。结果显示,与正常对照组相比,ASD和随后RS期间,包括收缩和延伸指数以及运动性指数在内的所有这些参数都显著且持续降低(图3B-G)。以上数据表明,在ASD和随后的RS期间,小胶质细胞突起的活动持续降低。
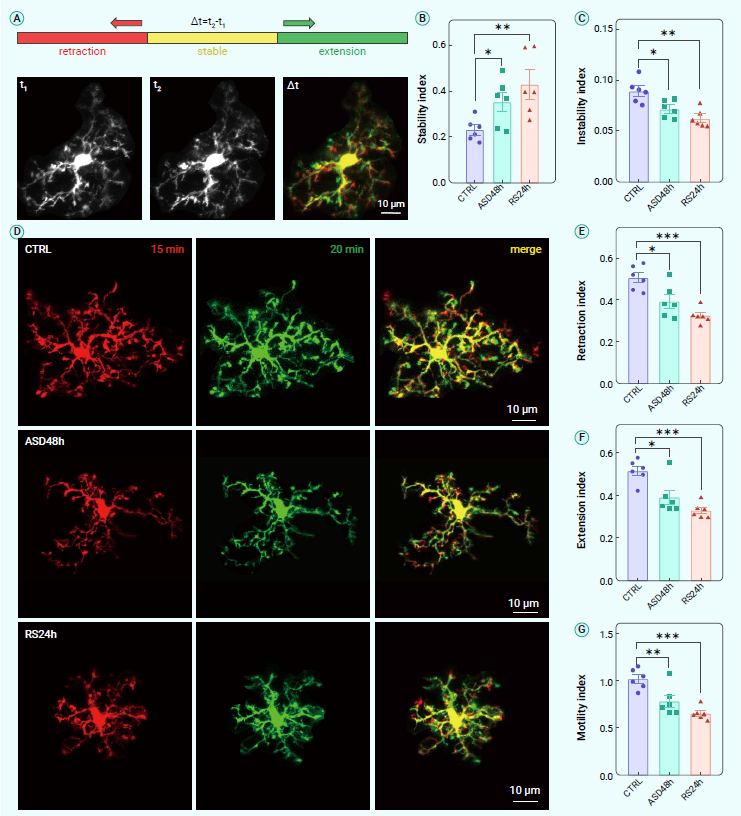
图3. ASD和随后RS期间小胶质细胞突起活动的体内成像
3.在ASD和RS期间,小胶质细胞的吞噬作用持续增强
作为中枢神经系统的巨噬细胞和突触修剪者,小胶质细胞通过其吞噬功能参与免疫防御以及突触的修剪和重塑。为了评估ASD和随后的RS期间小胶质细胞的吞噬活性,作者追踪了CD68的表达水平和定位(CD68参与单核细胞/巨噬细胞吞噬,清除细胞碎片和死亡细胞)。通过对小胶质细胞免疫荧光染色(图4A),结果发现,相比于正常对照组,ASD和RS组小鼠的不同大脑区域中CD68的表达持续增加(图4A-C)。因此,这些数据表明,在ASD和随后的RS期间,大脑区域中小胶质细胞的吞噬作用持续增强。
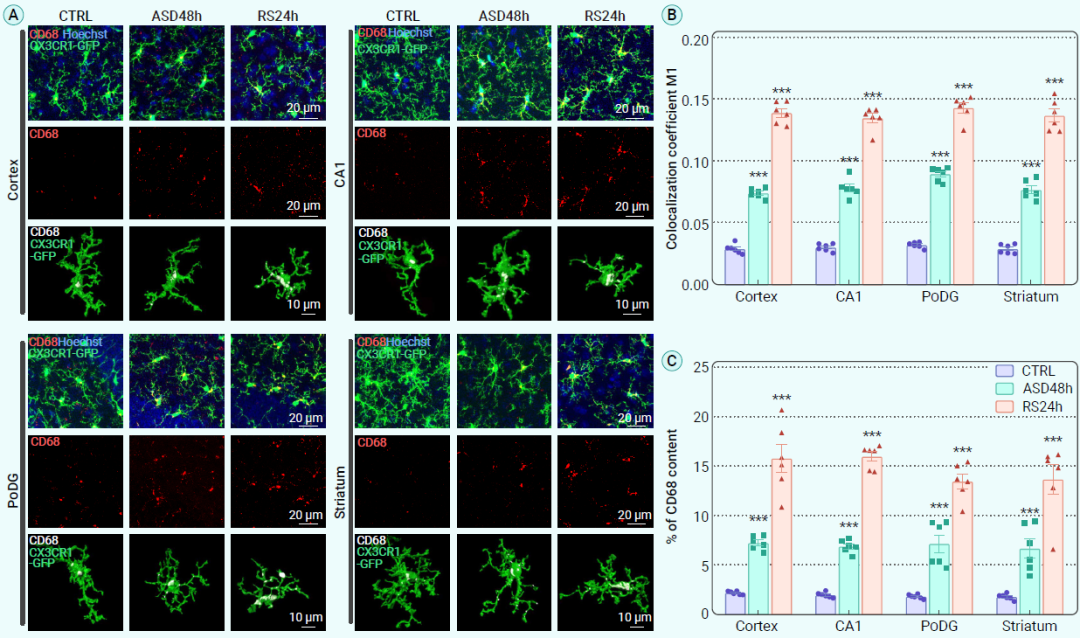
图4. ASD及RS后小胶质细胞的吞噬活性持续增强
4.在ASD和RS期间,促进了神经元树突棘的丢失
先前的研究表明,小胶质细胞的活动可以影响突触可塑性。为了研究ASD和随后RS对突触可塑性的影响,作者评估了整个ASD/RS过程的小鼠大脑躯体感觉皮层中树突棘的形成和丢失(图5A-B)。通过对同一神经元的树突结构进行动态追踪,研究发现,与正常对照组小鼠相比,ASD和RS组小鼠的神经元树突棘的形成率没有显著差异,但是树突棘的消除率显著增加(图5C-F)。因此,很可能是由于小胶质细胞激活持续处于低水平,导致在ASD和随后的RS期间过多的树突棘丢失。
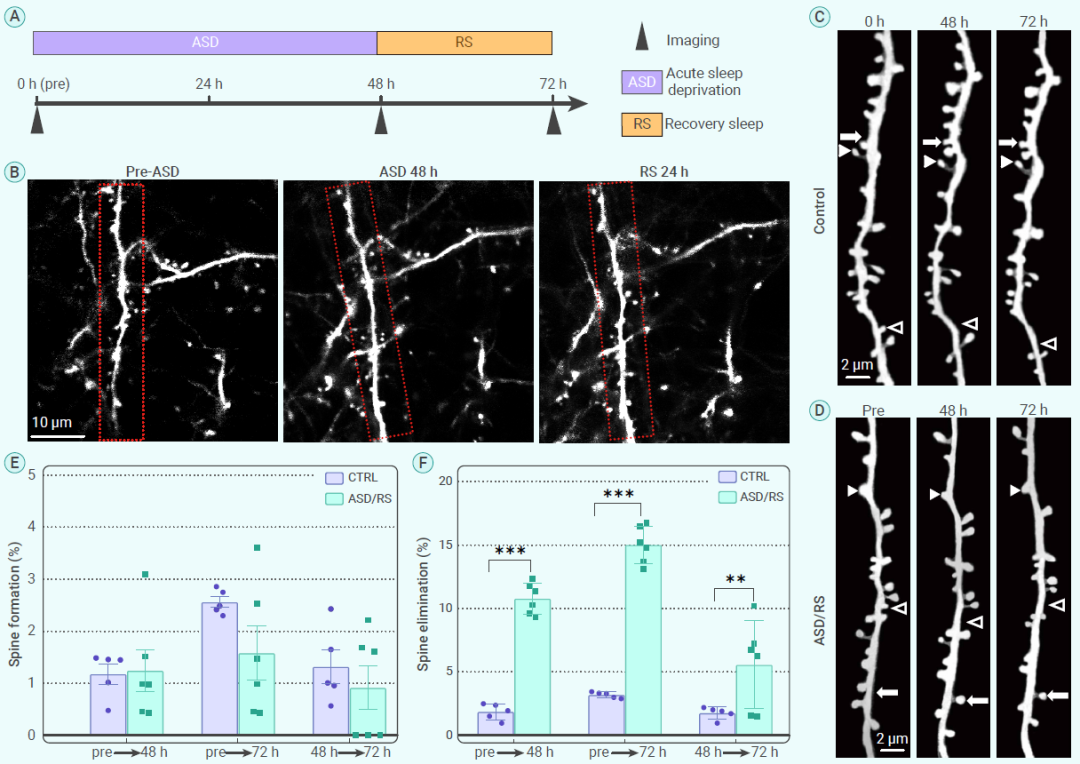
图5. ASD和随后的RS期间对树突棘的影响
5. 在ASD和RS期间,大脑的代谢水平受到影响
上述数据表明,在ASD和RS期间小胶质细胞的激活和神经可塑性可能会受到影响。为了进一步探索背后的分子机制,作者采用LC-MS/MS方法来评估它们对整个大脑代谢微环境的影响。热图显示,在ASD和RS之后,相当数量的代谢物水平被上调或下调,显著改变了小鼠整个大脑的代谢组(图6A-C)。基于模型的准确性和可靠性,作者接下来研究了大脑中对ASD和随后RS反应不同的代谢物。结果显示,与正常对照组相比,ASD后大量代谢物的水平显著改变,而RS后恢复了许多代谢物的水平(图6D-E)。此外,功能和通路富集分析的结果发现,大脑中最受影响的代谢途径为甘油磷脂代谢、烟酸和烟酰胺代谢等(图6H)。以上结果揭示了在ASD期间大脑的广泛代谢变化以及RS对代谢水平的恢复作用。
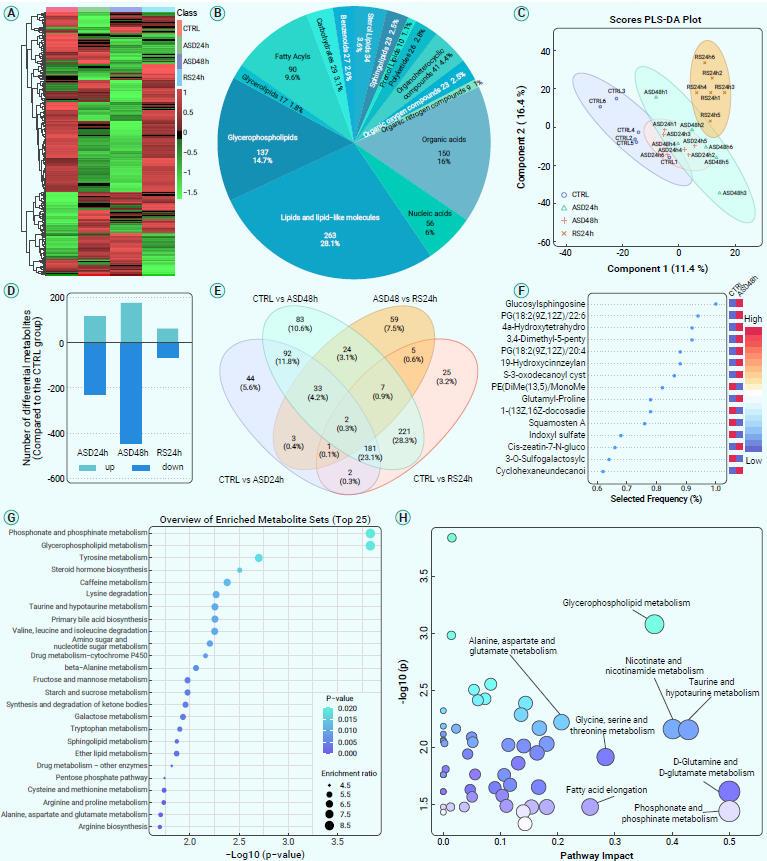
图6. ASD及RS期间的全脑代谢分析
6.部分代谢途径可能在短期睡眠恢复(RS)内无法恢
根据作者的代谢组数据,RS内大部分代谢物恢复到正常水平(图7A)。然而,即使在这个睡眠恢复期后,仍有许多代谢物没有恢复到正常水平。为了探究在RS期间观察到的小胶质细胞持续激活背后的机制,作者研究了那些丰度没有恢复的大脑代谢物。根据通路富集分析,在24小时睡眠恢复后,许多涉及甘油磷脂、牛磺酸和次牛磺酸等代谢物恢复到正常水平(图7B)。在不可恢复的代谢物中,包括甘油磷脂、磷酸盐和磷酸酯代谢在内的代谢途径显著富集(图7C)。接下来,这些可恢复和不可恢复的代谢物被导入Cytoscape,基于KEGG途径数据库可视化它们的相互联系(图7D和7E)。结果揭示了11个核心代谢物的主要途径,作者关注了其中与神经功能和疾病密切相关的不可恢复代谢物(图7F-M)。以上结果从RS中确定了可恢复和不可恢复的代谢物,从而揭示了可能与ASD和RS期间小胶质细胞持续激活和树突棘加速消除密切相关的关键代谢物和途径。
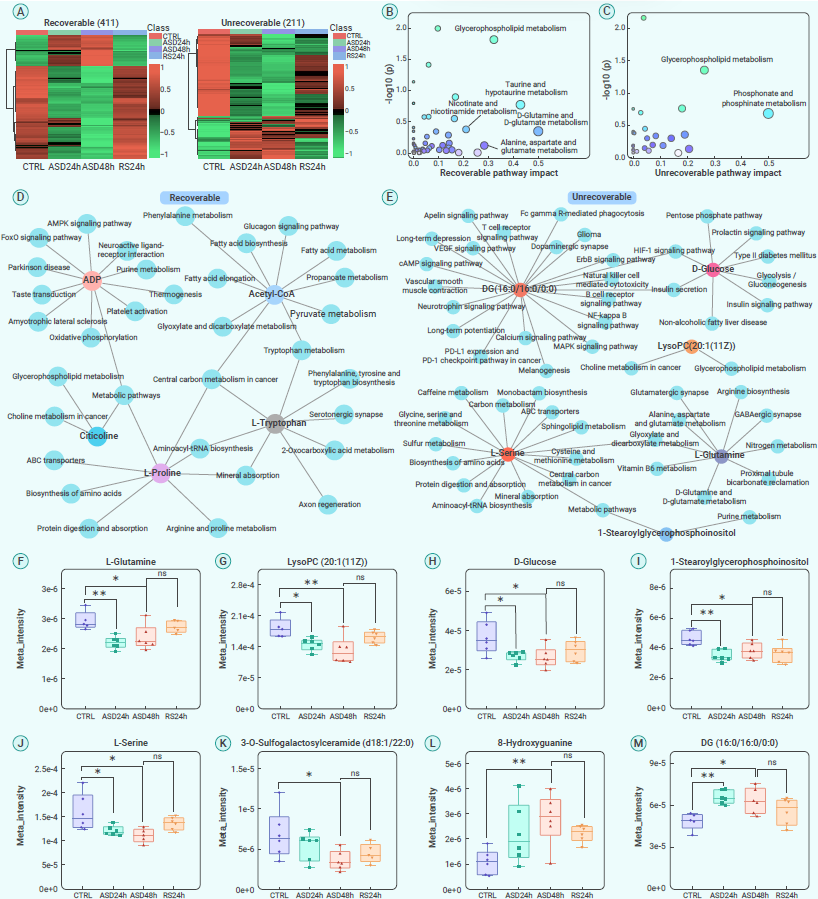
图7. 在RS 24小时内可恢复/不可恢复的代谢物和途径
结论
在这项研究中,作者建立小鼠急性睡眠剥夺(ASD)模型,通过使用多种转基因荧光小鼠模型和神经影像技术,并记录了在ASD和RS期间大脑区域中小胶质细胞突起活动性降低,吞噬作用增强的持续变化以及整个过程中树突棘的加速消除。随后,基于 大脑组织代谢分析,发现在ASD和RS期间观察到的持续小胶质细胞激活和树突棘加速消除的潜在机制。这项工作可能有助于阐明不规律睡眠导致神经系统疾病的细胞和分子机制。
在该研究中,研究人员使用了赛昂斯睡眠剥夺仪(SANS,型号SA109)来进行实验操作。

睡眠剥夺仪(SA109)
SA109型大小鼠睡眠剥夺仪参考进口同类产品重新升级设计,提供一个标准的居住环境,其中包括食物和水的支持。它不需要动物一直处于行走的状态,通过可调节螺旋杆的旋转周期,实现动物睡眠剥夺试验;系统还配备了昼夜节律灯光控制系统,预留额外接口,可配合光遗传实验、钙离子成像实验用。

产品特点
1、旋转模式:正转,反转,交替,随机;
2、间歇时间:0-240s,步进为1s;
3、周期范围:2s~40s,步进为1s,周期范围内任意编程周期时间段;
4、预约功能:如定时启动为凌晨3点,运行5小时,上午8点停止,每天可以重复,期间无
需人工值守,自动运行和停止;
5、测试时长:持续测试时长为1-900小时;
6、静音模式:实验过程中无任何机械声音,静音标准低于于40分贝;
7、带有昼夜节律灯光控制设置,自定义设定昼夜节律灯,获得更好的睡眠剥夺效果;
8、具有外部触发接口,支持钙离子成像,光遗传,脑电设备。