大家都在搜

|
在进行分子表达操控中,经常听到或使用到的一个技术便是Cre/Loxp或Flp/FRT重组酶系统。简单来说,这类重组酶系统是通过特异位点重组酶(Site-specific recombinases, SSRs)介导重组酶特异识别位点(Recombination tar-get sites, RTs)间的重组,来实现特异位点的基因敲除、基因插入、基因翻转和基因易位等操作。由于该技术能够有效克服其他类型重组技术的非特异性或重组效率低等缺点,近年来已逐渐在功能基因研究领域占据了主导地位。 Cre-loxP的基本原理 Cre(Cyclization Recombination Enzyme)是一种重组酶,来源于P1噬菌体,其基因编码区序列全长1029bp,为38kDa大小的、由343个氨基酸组成的多肽单体蛋白。Cre重组酶的C-末端结构域包含催化活性位点,能够催化DNA分子中特定位点之间的重组,同时,Cre还能识别特异的DNA序列,即loxP位点,使两个loxP位点间发生基因重组。
Cre/loxP诱导基因重组的方式 一般而言,当细胞基因组内存在两个LoxP位点时,Cre重组酶会诱导两个LoxP位点间的序列发生重组。重组的结果取决于两个loxP位点的方向,主要有以下几种可能:
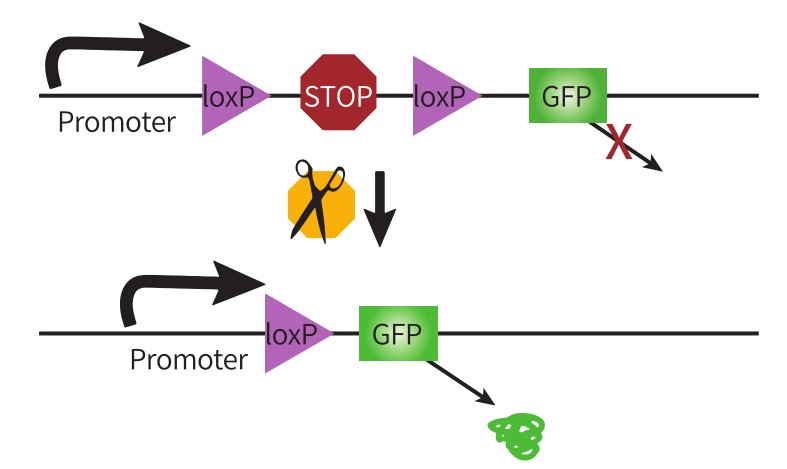
01 LSL序列(条件性基因选择) 将LoxP2和转录终止信号盒(Transcription STOP cassette)插入启动子和目的基因之间,转录终止信号盒两端各有一个同向的LoxP位点,组成LoxP-STOP-LoxP-gene模式,即LSL序列。
图2 Cre依赖的基因表达-LSL策略
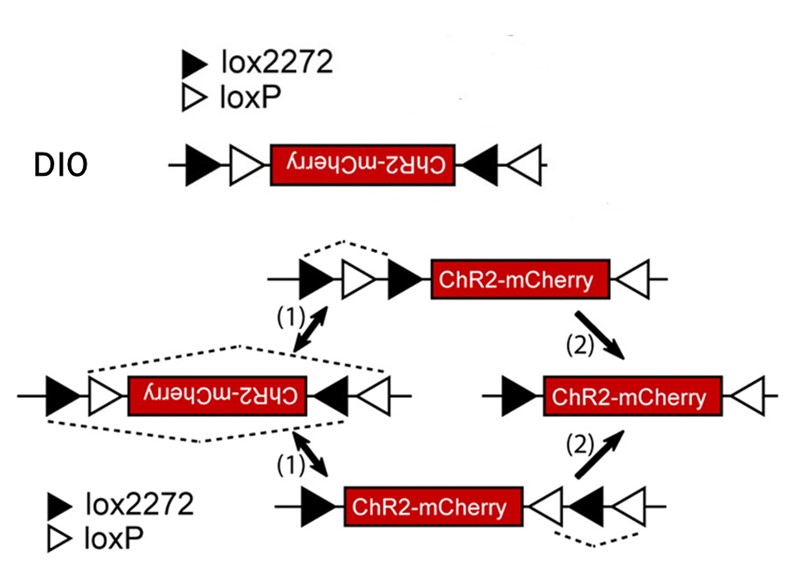
02 DIO/DO序列 通过引入两对不相容的反向Lox位点LoxP和Lox2272,经过两组Lox位点的两轮重组可达到一种稳定状态。也就是可以通过Cre重组酶的存在与否来控制基因的表达。
图3借助LoxP和Lox2272的FLEX策略实现Cre依赖的基因表达
03 MADM系统 双标记嵌合体分析MADM(mosaic analysis with the double markers)系统,是基于Cre诱导发生Translocation过程实现的。通过同源重组将两个相互嵌合的标记基因(荧光蛋白)分别定位到同源染色体上的相同位点,位于同一条DNA链上的荧光蛋白N端(C端)和另一种荧光蛋白N端(C端)之间插入一个Loxp位点。在没有Cre的情况下,不表达功能性的GFP或RFP;但在特定时期、特定细胞中表达Cre时,Cre能诱导带有两种不同荧光蛋白N端的DNA链与另一条带有相同荧光蛋白C端的DNA链在细胞有丝分裂G2期发生重组,使得两个子代细胞分别表达有任一种有功能性的荧光蛋白(称为:X-segregation),或者其中一个子代细胞表达两种荧光蛋白而另一种子代细胞不带任何功能性荧光蛋白(称为:Z-segregation)。另外,重组也可能发生在细胞周期的G1期或者G0期,在这种情况下,细胞同时表达两种荧光蛋白。
图4 MADM系统
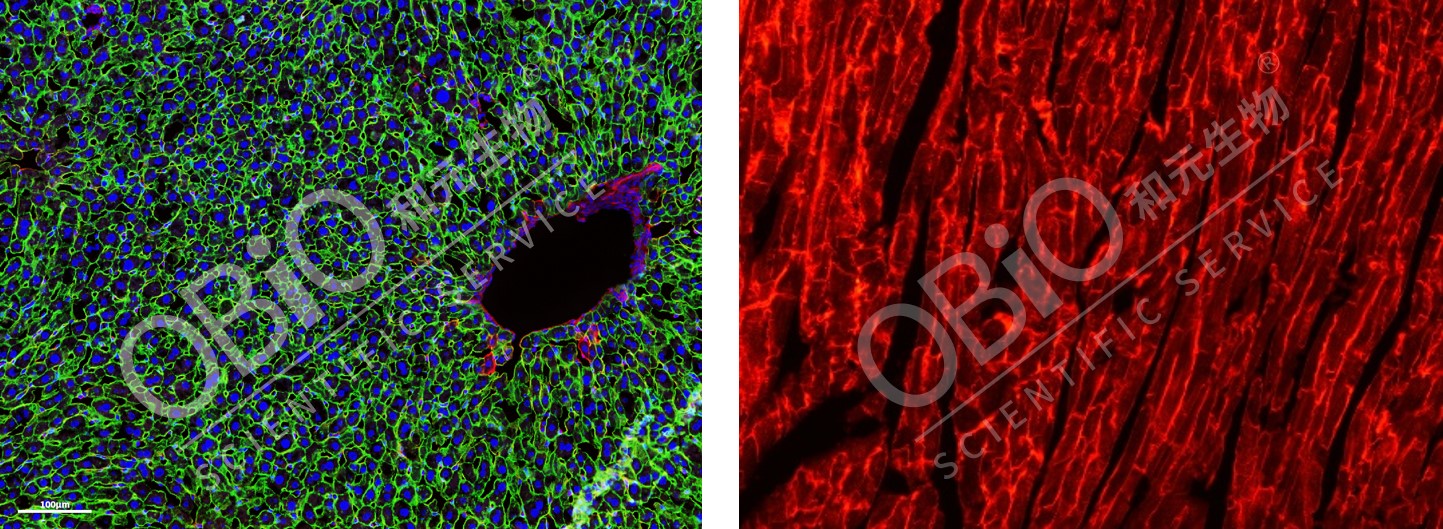
Cre/loxp系统的应用 通过病毒引入Cre或loxP元件,结合一种转基因小鼠可达到对某类细胞进行特异性标记或基因操作的目的。 LSL策略应用 mT/mG(ROSA-pCA-loxp-mT-pA-loxp-mG-pA)鼠是一种双荧光报告鼠,这种鼠在正常情况下表达定位在膜上的红色荧光蛋白(TdTomato),而当细胞表达Cre后,则表达定位在膜上的绿色荧光蛋白(GFP)。
图5肝脏组织(左)、心脏组织(右)
管吉松教授团队将AAV-hSyn-Cre注射于 Ash1lfl/fl小鼠右侧AUD区条件性敲除Ash1l基因,发现Ash1l以细胞自主效应的方式介导皮层神经元的活动依赖的突触修剪过程。(点击阅读:Neuron | 拯救星星之子!上科大管吉松组揭示ASH1L单倍不足致小鼠自闭症的神经机制)。
图6 AAV-Cre注射Flox小鼠实现条件性敲除
DIO策略应用 Thy1是锥体神经元的特异性启动子,在前脑、海马、杏仁核、丘脑及视网膜等区域均有丰富的表达, Thy1-cre鼠配合rAAV载体可在以上脑区实现细胞特异性标记、光遗传学和化学遗传学操纵、在体钙成像记录,或者组织特异性的基因编辑。
图7 AAV-DIO注射Thy1-Cre鼠实现特异性标记
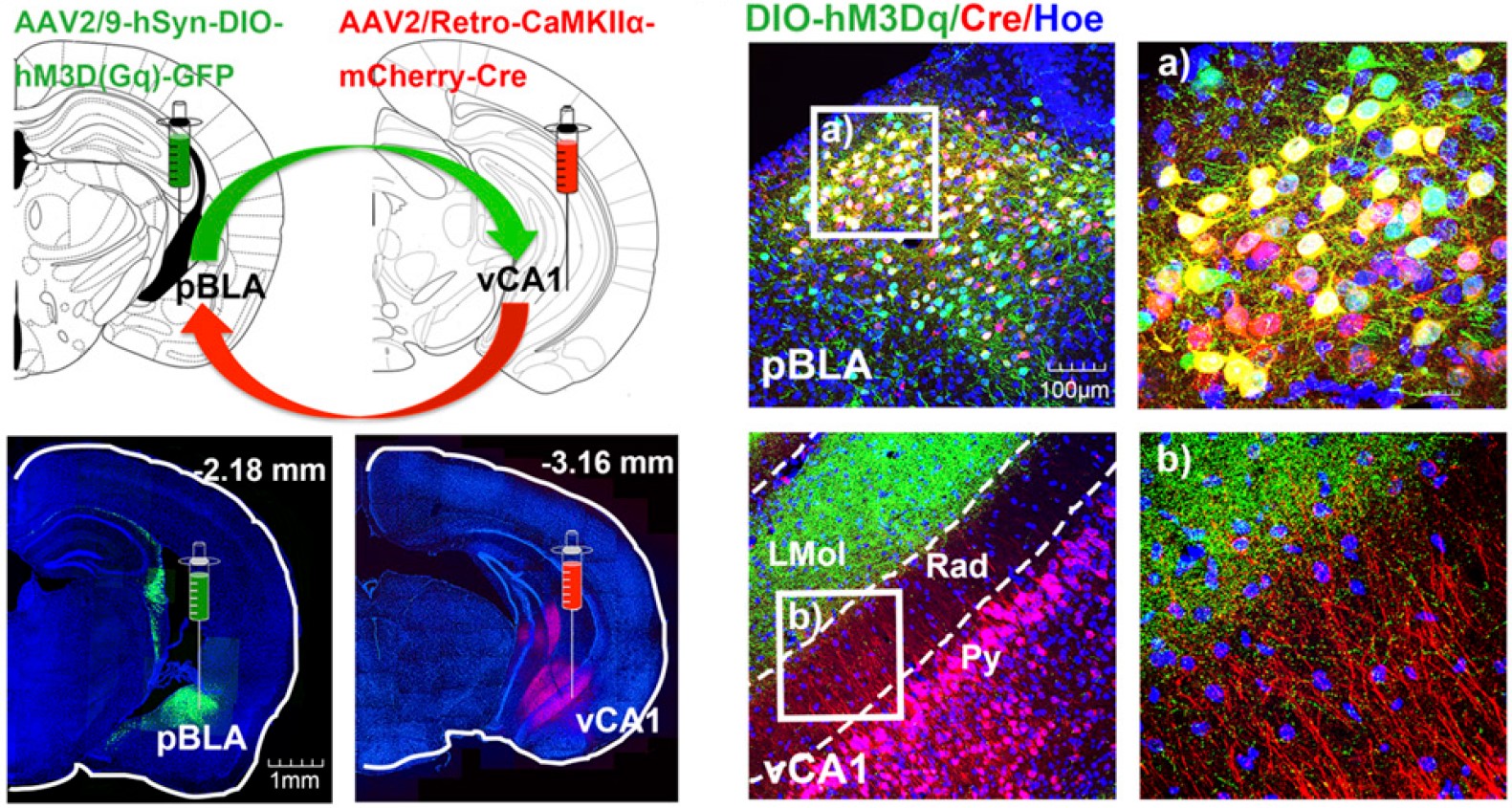
运用神经示踪技术对大脑特定神经环路的结构和功能进行解析时,亦可以借助Cre重组酶系统和AAV血清型(比如rAAV2/9、rAAV2/retro、rAAV2/1)结合适用,达到特异性对神经环路标记和功能研究的目的。
图8 Cre-DIO系统结合病毒示踪技术实现对特异环路标记及调控
MADM系统应用 借助心肌细胞特异性启动子Tnnt2驱动cre酶的表达,配合MADM-ML-11TG/GT鼠追踪心肌细胞细胞分裂的状态。(点击阅读:【Cell Stem Cell】鸡尾酒疗法促进心肌细胞增殖和心脏再生)。
图9 MADM系统应用
Cre病毒列表(部分)
改造的Cre/loxP系统 经过现代基因工程方法对Cre和loxP元件的改造,Cre/loxP系统实现了更加丰富的条件性重组策略。
图10 tamoxifen诱导的Cre-ER系统
图11 FISC体系
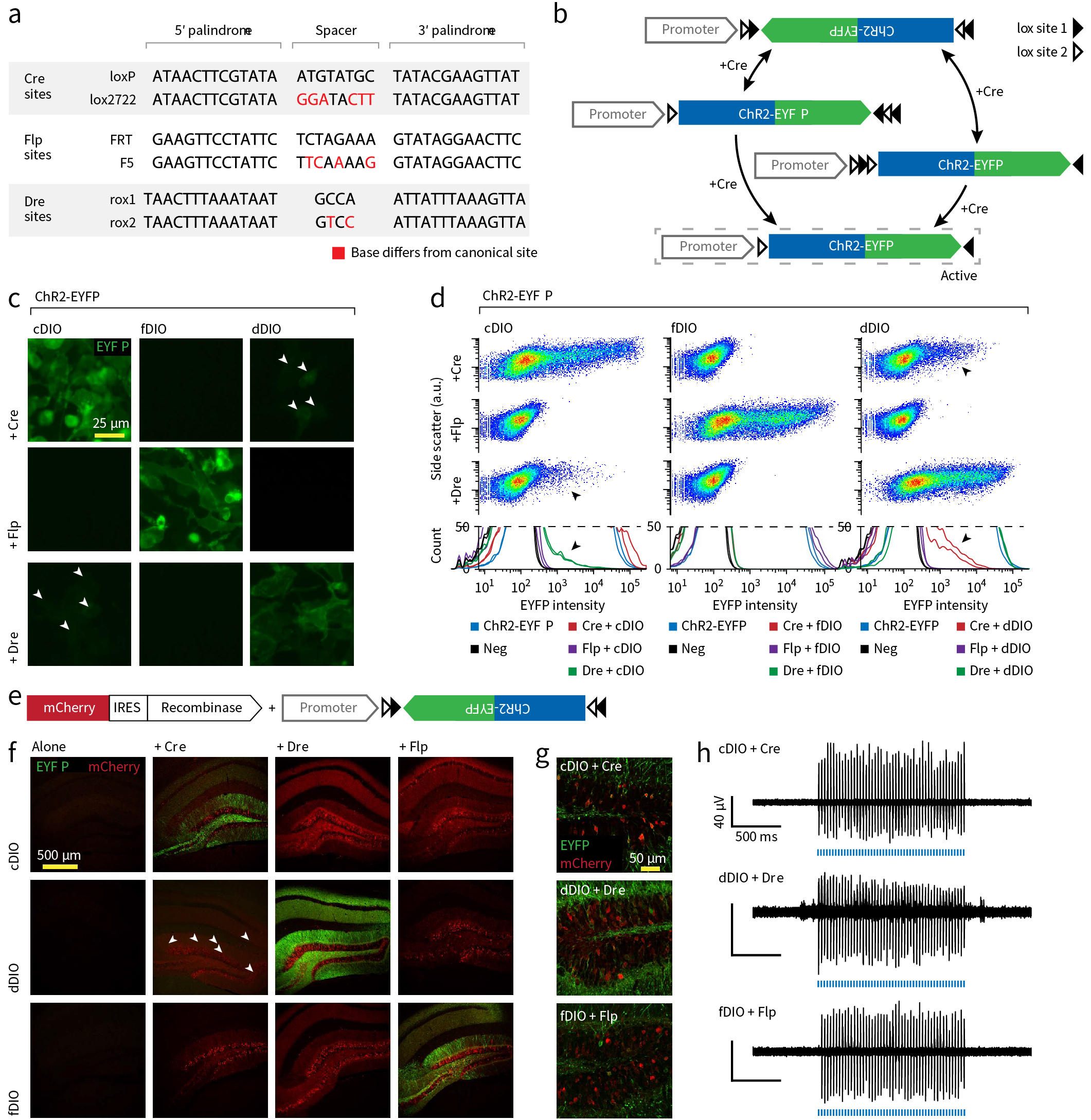
其他重组酶系统 除Cre-LoxP系统外,类似的还有与Cre-LoxP系统无交叉影响的vCre-vLoxP、sCre-sLoxP系统,及Flp-FRT/F5系统和Dre-Rox1/Rox2系统,对应的表达方案分别为fDIO和dDIO系统。多套重组酶体系的存在为研究中设计多个限制条件提供了便利,灵活应用Cre、Flp、Dre重组酶系统将有助于更深入课题的开展(了解更多重组酶内容,点击阅读:【知识分享】一半海水、一半火焰,分子表达操控中高频出现的Cre、Flp、Dre等重组酶系统如何工作)。
图12 Cre\Flp\Dre系统比较
[1] Front Genet. 2016 Feb 19;7:19. doi: 10.3389/fgene.2016.00019. [2] Sci Adv. 2019 Feb 20;5(2):eaat3210. doi: 10.1126/sciadv.aat3210. [3] Nat Methods. 2014 Jul;11(7):763-72. doi: 10.1038/nmeth.2996. [4] J Biol Chem. 2020 Jan 17;295(3):690-700. doi: 10.1074/jbc.RA119.011349. [5] J Neurosci. 2008 Jul 9; 28(28): 7025–7030.doi: 10.1523/JNEUROSCI.1954-08.2008 [6] Lab Anim Res. 2018 Dec;34(4):147-159. doi: 10.5625/lar.2018.34.4.147. [7] Nucleic Acids Res. 2004 Nov 18;32(20):6086-95. doi: 10.1093/nar/gkh941. [8] Dis Model Mech. Sep-Oct 2009;2(9-10):508-15. doi: 10.1242/dmm.003087. [9] Nucleic Acids Res. 2011 Apr;39(8):e49. doi: 10.1093/nar/gkq1280. [10]Cell. 2005 May 6;121(3):479-92.doi: 10.1016/j.cell.2005.02.012. [11] Nat Commun. 2020; 11: 3708. doi: 10.1038/s41467-020-17530-9
|

更有优质直播、研选好物、福利活动等你来!