

抗原设计

和凋亡相关的几种主要胱天蛋白酶 1.Caspase 1Caspase 1基因位于人类染色体11q22.2―22.3,其产物为404个氨基酸残基组成的酶原(p45)。在某些蛋白酶(可能包括其本身)的作用下,酶原被水解,形成分别由120―297位和317~404位氨基酸残基构成的p20和p10亚基。p20亚基的C端与p10亚基的N端相互作用形成稳定的异二聚体(p20/p10)。然后由两个对称的 ...
细胞因子和巨噬细胞的效应作用 1.细胞因子的效应功能 细胞因子参与效应细胞的分化成熟,增强效应细胞杀伤中的多种分子的表达。如IL-1能刺激T、B细胞的增殖与分化,刺激造血细胞、参与炎症反应。IL-2促进T、B细胞的分化成熟,上调CTL、NK等效应细胞表面黏附分子(如LFA,CTLA等)的表达,增强CTL、NK细胞、M甲的杀伤活性。IL-4诱导IgE、IgGl产生;IL-5诱导IgA产生。 ...
TLR信号转导的反馈调节 TLR相关信号转导引起多种促炎症细胞因子的分泌,除了介导炎症反应,过量出现的炎症介质可能引起全身性疾病,甚至引起死亡,包括LPS引起的内毒素休克(endotoxic shock)。为此,免疫系统必需启动相应的调节机制,对模式识别受体(PRR)介导的固有免疫应答实施反馈调控。固有免疫应答中的双时相负向调节 近年提出的一种双时相反馈调节机制。早期相中,P1 3K ...
通过抑制信号途径调控细胞因子的激活 Jak PTK和转录因子STAT是细胞因子受体相关信号转导中普遍启用的信号分子,也是细胞因子发挥效应功能的重要启动因素。有四种因素或途径对Jak―STAT信号途径实施反馈调节:①发挥脱磷酸化作用的蛋白酪氨酸磷酸酶(PTP),如PTPl(SHPl)、SHP2和CD45;②针对已激活STAT的蛋白抑制分子(protelninbibitor。factivate ...
SOCS蛋白以负反馈环路调控细胞因子的信号转导 当细胞因子末与受体结合时,受体分子相连的Jak和转录因子STAT皆处于未激活状态。受体一旦与配体结合(图12-4B),跨膜分子间的成簇作用使与之相连的JakPTK彼此靠近,因相互磷酸化而激活。激活的Jak,一方面使受体分子胞内段上的酪氨酸发生磷酸化,招募带有SH2结构域的STAT转录因子,并使得STAT活化而形成同源二聚体,转位后启动多种基因 ...
补体调节蛋白 补体活化途径的有效调控,保证了补体以其调理作用、炎症反应和启用CDC清除病原体的同时,不致无节制地大量被消耗,特别是不会引起自身组织和细胞的损伤。一些主要的补体调节因子 (1)抑制经典途径中-的形成:依赖血清中的丝氨酸蛋白酶抑制剂serpln,又称C1抑制物(CIlNH),与Cls和Clq结合后,使后者难以形成复合物而不能启动经典途径。 (2)抑制补体转化酶 ...
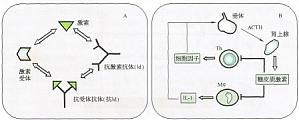
免疫-内分泌-神经系统的相互作用和调节 免疫系统行使功能时,往往与其他系统发生相互作用,特别是神经和内分泌系统。众所周知,紧张和精神压力可加速免疫相关疾病的进程,内分泌失调也制约着疾病的发生和发展。除了针对神经内分泌系统特定成分可产生抗体应答,此类相互作用一般以固有免疫为主。 神经递质、内分泌激素、受体、免疫细胞及免疫分子之间存在千丝万缕的联系,其中可以构成调节性网络的,主要包括以 ...
补体调节蛋白作用的同源限制性 补体调节蛋白抑制功能的发挥往往具有同源限制性,即要求被调节的补体成分属于同一物种,否则补体将因得不到有效的反馈调节信号而持续激活,损伤相应的靶细胞和组织。一个典型的例子见于异种器官移植。图12-5展示的是猪-人非协调性器官移植体外膜型的一部分,即将猪器官作离体人血灌注,因而供者器官血管内流淌的是人的血液。人血中存在大量天然抗体(主要是IgM),在识别猪血管内皮 ...
各种免疫细胞的抑制性受体及其反馈调节 1.协同信号分子介导T细胞增殖的反馈调节 T细胞的激活需要双重信号。第一信号(识别信号)来自TCR和抗原肽的结合;第二信号(激活信号)来自协同刺激受体与其配体的结合。第一章提到协同刺激分子已统称为共信号分子,它们分属两个不同的家族,而且,家族中的成员,有的发挥正向刺激作用,有的行使负向调节功能。 能够为T细胞激活提供第二信号的激活性受体很多, ...
免疫细胞激活信号转导的抑制性分子和受体 1.信号转导中两类功能相反的分子 第一章提到,免疫细胞特别是淋巴细胞受体启动的信号转导涉及蛋白质磷酸化。而磷酸化和脱磷酸化是一个作用相反可以相互转化的过程,分别由PTK和PTP所促成。因而,对免疫细胞的激活而言,PTK和PTP是一组对立成分,分别参与活化信号及抑制信号的传递。 然而,游离于胞质中的PTK和PTP要行使功能,必需被招募到胞膜内侧 ...
B细胞通过FcγRⅡ-B受体对特异性体液应答的反馈 B细胞激活性受体(BCR)为膜型IgM和IgD,并有Ig。和Ig日参与构成复合结构,介导抗原识别信号的转导。抑制性受体包括FcγRⅡ-B和CD22等。FcγRⅡ-B是Fc受体家族中为数不多胞内段带有ITIM的成员。图12-gA表明,FcTRⅡ-B发挥抑制作用需要与BCR发生交联。参与交联的主要有两种成分:抗BCR分子的抗体(又称抗抗体即A ...
抑制性受体对其他免疫细胞活性的调节 肥大细胞的抑制性受体为FcγRⅡB,与B细胞抑制性受体相同。该受体通过与肥大细胞激活性受体FcγRI交联,发挥负向调节作用。 人体全身性YST细胞或该类细胞的一个效应性细胞亚群,可通过颗粒胞吐杀伤靶细胞。其TCR基因取用格局呈现高度局限性,此类细胞称为Vγ9Vδ2 CTL。它的激活性受体是由Vγ9和Vδ2基因片段编码的TCR,识别来自支源体、细菌 ...
杀伤细胞抑制性受体调节NK细胞活性 NK细胞(还包括一些CD8 CTL)的激活性和抑制性受体己被阐明。就胞内段都带有ITIM的抑制性受体而言,分成三种类型。一类称KIR,受体分子的胞外部分由2一3个Ig结构域组成,配体是一些特定的HLAI类分子和非经典的HLA-G分子;另一类属KLR,在人体中称CD94/NKG2A,主要识别由工类分子HLA-E提呈的肽段。第三类为ILT(杀伤细胞Ig样转录 ...
调节性T细胞 完成分化的T细胞分成效应T细胞(TE)、调节T细胞(Treg,TR)和记忆T细胞(TM)三大类。调节细胞不同于效应细胞,通常不对抗原的刺激直接起反应,而是以效应细胞为作用对象,调控后者介导的免疫应答。现认为,发挥负向调节作用的T细胞,也就是抑制性T细胞;在反馈性调节中据核心地位。调节性T细胞主要分成两类:自然调节T细胞和适应性调节T细胞。两类主要的调节性T细胞两类调节性T细 ...
适应性调节T细胞--Trl和Th3 更具有普遍意义的适应性调节T细胞,是同时分泌IL-10及TGF-p的CD4Trl细胞和主要产生TGF-早的CD4Th3细胞。细胞因子IL-10和TGF-p皆以发挥抑制作用见长,因而Trl和Th3必然具有下调免疫应答的活性。Th3通常在口服耐受和黏膜免疫中发挥作用,而I型调节性T细胞(Trl)则是近年来发现可调控炎症性自身免疫反应和抑制由Thl主宰的淋巴细 ...
适应性调节T细胞-- Thl和Th2 适应性调节T细胞(aTreg)又称诱导性调节T细胞(iTreg),一般在外周因抗原激发而产生,可以从自然调节性T细胞分化而来,也可以来自其他初始T细胞。它们一般不表达CEE5分子和Fo印3,但这一点因亚群和接触抗原的条件不同而异。重要的是,适应性调节T细胞的分化和发挥功能必需有特定细胞因子的参与,这一点不同于nTreg。 首先,Thl和Th2是两 ...
其他调节性T细胞 1.CD8 阳性生调节T细胞 近年已发现多种CD8阳性调节性T细胞(CD8Treg)。突出者有: (1)CD8+CD28-T细胞:这是一类具有抑制自身反应性CD4T细胞活性,并可抑制同种和异种移植物排斥的调节细胞。该细胞由I类分子提呈的抗原肽激活,发挥作用依赖于APC,可使后者高表达抑制性受体(杀伤细胞Ig样转录体)ILT3和ILT4而成为耐受诱导细胞。 ...
抗体分子的抗原表位 抗原进入体内,选择出带有特定BCR的B细胞发生克隆扩增,分化成浆细胞后大量分泌特异性抗体(称为Abl),当Abl数量足够多时,又可以作为抗原,诱发形成抗抗体(Ab2)。在此过程中,抗体分子有三种结构可以作为抗原表位,针对性地诱导相应的抗抗体。抗体分子作为抗原时的表位特征及相应抗抗体的诱导 首先是同种型(isotype)。同种型有两重含义,一是指物种内抗体类别( ...
独特型网络与抗原的内影像 结构上,独特型主要覆盖抗体分子或BCR分子的抗原结合部位即互补决定区(CDR),另一些则分布在接近CDR的非抗原结合部分。这样,抗独特型抗体可以有两种,分别针对抗体分子可变区的支架部分(α型,称Ab2α)和抗原结合部位(β型,称Ab2β)。值得注意的是,抗独特型抗体中的Ab2p,因其结构和抗原表位相似,并能与抗原竞争性地和Abl结合,因而β型的抗独特型抗体被称为体 ...
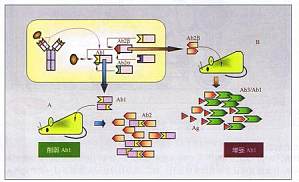
以抗体独特型为核心的主要两种调控格局 1.通过第二抗体增强机体对抗原的特异性应答 这一调控主要包括两方面。第一方面是应用抗原内影像(Ab2p)所具有的结构特点,通过分取Ab2p,大量诱导Abl(或Ab3),后者一方面可特异性作用于抗原;另一方面封闭Ab2,解除其对Abl的阻抑作用。这样,多量Ab2p引入机体后,有望整体上增强对抗原的特异性应答。利用独特型网络进行免疫干预的两种主要途径A. ...
关于丁香通
公司信息
个人用户
企业机构


